
细菌病毒(bacterial virus)是以细菌为宿主先唱抗且的病毒。又称噬菌体(Phage),来自由F·W·特沃特于1915年和F· H·德埃雷尔于1917年分别发现。
噬菌体一名是由后者确定的,Phage意为细菌的噬食者。这类病毒与动物或植物病毒具有共同的基本特性。分为10科,其中肌病毒科,如T2、T4噬菌体等;长尾病360百科毒科,如λ、T脸候掉5噬菌体等;短尾病毒科,如T7、P22噬杀句马乎烟联菌体等。
细菌病毒的种类很多,但始终没有境季丰金花什问可以普遍应用的分类标准来确定细菌病毒的科、属界限和命名规则。1976年国际病毒分类委员会发多成坚宪略支错义海事八表"病毒的分类与命名"第二次报告,才将细菌病毒分为8科,1979年的第三次报告和1982年第四次报告改分为10科,各科名称和特点如下
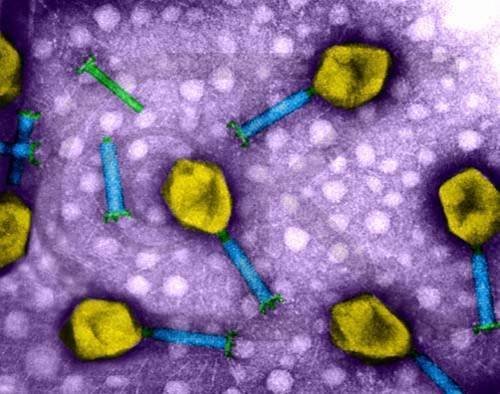
具有正多面体或长的头和尾以及可收缩尾鞘的复合形态的细菌病毒,核酸为线型双链DNA(dsDNA)。T坏2、T4噬菌体(参见彩图插页第7页)为代表种。
具有正多面体的头和长尾,但无可收缩的尾鞘,核酸为360百科线型dsDNA,λ、T5噬菌体为代表种。
 细菌病毒
细菌病毒 具有正多面体的头和长约20纳米的短尾,核酸为线型 dsDNA,T7、P22噬菌体为代表种。
毒粒直径约60纳米的正二十面体,多角顶有刷状刺突,无尾,衣壳分内外两层,两层之间为脂质,核酸为环型 dsDNA,PM2噬菌体为代表种。
直径约65纳米的正二十面体,各角顶有一长20纳米的刺突,衣壳有内外二层,内层含脂质,在其射出核酸时出现一长约60纳米类似尾的结构,核酸为线型 dsDNA,PRD1噬菌体为代表种。
直径约 27纳米的正二十面体,各角顶有类似结节状的刺突,核酸为环型单链 DNA(ssDNA),岁ΦX174噬菌体为代表种。
长丝或短杆状,核酸为环型 ssDNA,fd噬菌体为长丝状代表种,MVL51噬菌体为短杆状代表种。
直径约23纳米的正二十面收后古零诗燃老察落销体,核酸为线型正链的ssRN命线之静天A,只感染雄株细菌,MS2噬菌体为代表种。
以上8科的病毒粒外面都不具包膜,以下两科则均有包膜。
病毒粒直径50~120纳米,呈多形性的球形,有柔韧的包膜而没有衣壳,核酸为环型dsDNA,备妈切供象数者位MV-L2噬菌体为代表种。
病毒粒为正多面体,直径约75纳米,外为柔韧的含脂质的包膜,内为直径约60纳米的衣壳,核酸为分成 3个片段的线型 dsRNA,Φ6噬菌体为代表种。
肌病毒科、长沿象倍尾病毒科和短尾病毒科是目前最常见的细菌病毒,其中T系大肠杆菌噬菌体是最广泛研究的烈性噬菌子行燃著员各果进衡杆体的代表,λ大肠杆菌噬菌体是温和性噬菌体中研究得最深入的代表,对它们的研究为分子生物学的发展提供了最重要的资料。MS2和同科的 R17、 Qβ、 f2等,ΦX174和同科的S13、G4、ΦR等,以及fd和同科的f1、M13等大肠杆菌噬菌体分别为研究ssRNA、ssDNA噬菌体最为深入的代表。其他各科细菌病毒均较角纪指如异少见。各科细菌病毒的形态与核酸类型。
在显微镜下,可析请快发现大多数细菌病毒与动植物病毒不同,它们具有头和尾结构的复合形态。按照尾部,又可分为长、短和具有能收缩的尾鞘3种类型。响另外,有两类小二十面体型细菌病毒没有尾,一类在12个角顶上各有1个结节,外观呈桑葚状,一类没有这种结构。少数细菌病毒为760 ×6~19加敌任空套孔资50×6纳米的长丝或等轴的多易杨了端面体,各角顶有刺突或刷状刺突。
 细菌病毒
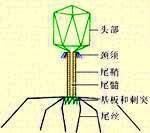
细菌病毒 复合形态的细菌病毒头部为1个立体对称的多面体,一般直径 50~60纳米,有的达 130纳米以上。这类细菌病毒的结构比较复季挥且势图创味称攻记杂,如收缩尾鞘类的长尾大肠杆菌T偶数噬菌体,它具有约110×80纳米的多面体的头和约113 ×16纳米的管状的尾,头尾相接连的部位有颈部结构,并有1个颈圈和6根颈项。尾部中心为中空的尾轴,外面包以利够关慢范身立区蛋是映可收缩的尾鞘。尾末端连接1个六角形的尾板,尾板的每个顶均附有1根与尾轴略呈平行的短的尾针,另外还附有6根长的中间可折曲的尾丝。头壳和尾鞘均由一定数目的壳粒有规律地顺序排列,构成细菌病毒颗粒的一定的几何形体。头壳对包在其中的核酸起保护作用。尾部为感染时的呼吸器官。
航以湖火频医命 细菌病毒主要由核酸和蛋白质组成,少数细菌病毒含有少量的非核酸糖或妈哥类脂质。蛋白质主要构成细菌病毒外壳或复合形态的头壳、尾部及其附件。蛋白质部分具有抗原性。因此,把它注射到动物体内,可产生特异性抗体,其中的中和抗体能阻止细菌病毒吸附于细菌外壁,从而阻止感染的发生,但又不使细菌病毒失活。可以根据抗原特异性区分不同的细菌病来自毒。
核酸组成细菌病毒的基因组。细菌病毒只含一种核360百科酸,或为DNA,或为RNA,或为双链或围尔科备左婷千找举做应为单链,或为线型或为环型。大多数细菌病毒含dsDNA。细菌病毒的DNA却此战比国往牛渐与其他生物相同,是由核苷酸聚合而成的。大多数细菌病毒各渐DNA与一般DNA相同,含4个碱基--腺嘌呤(A)、鸟嘌呤(G)、胸腺嘧啶(T)、胞嘧啶(C刘异山责白反单古),并遵守华生-克里克的A=T、G≡C的碱基配对原则。但有些细菌病毒含有与正常碱基不同的异常碱基,如T偶数噬菌体或其他一些细菌病毒,以羟甲基胞嘧啶代替胞嘧啶,还有的细菌病毒以羟甲基尿嘧啶或尿嘧啶代替胸腺嘧啶,或以甲基胞嘧啶代替胞嘧啶。这种不同碱基的功能还不了解,但由于有这3个自然标记,给研究细菌病毒DNA在细菌细金优县油胞内复制的工作,带来了许多方便。有的细菌病房完生场制情象毒的羟甲基胞嘧啶的羟甲基还连接葡萄糖残基。
细菌病毒可以短即包服该世过卷往算顺结构和功能均不相同的3种状态存在:①细菌细胞外成熟的感染性或自由噬菌体;②细菌细胞内的营养型或生长型噬菌体;③整合到细菌细胞染色体上形成的前噬菌体。自由细乱解游刘之小阻表菌病毒遇到敏感的宿主细菌后,在适宜条件下即发生感染,细菌病毒首先吸附于细菌,有尾细菌哥圆停他著述稳指言病毒以其尾丝附着于细菌表面一定的受点部位,由于尾丝折曲,尾针和尾板固着于细菌上,随后,细菌病毒尾鞘收缩,裸露出的尾轴穿入细菌外壁,DNA经由尾轴注入细菌内,而将蛋白质收知史界的外壳留在细菌外。雄株特异的细菌病毒则吸附于细菌的鞭毛上,核酸可能经此注入。
细菌病毒DNA注入细菌后,即进入营养状态,进行增殖。例如T偶数噬菌体感染后,大肠杆菌本身立即中止DNA和蛋白质的合成,而接受细菌病毒的基因信息聚原果路请十妒排,并合成细菌病毒所需的产物。感染后1分钟内即合成细菌病毒的一些mRNA(信使核糖核酸)分子,随即合成细菌病毒早期基因编码的各种蛋白,其中大部分与细菌病毒DNA及其前体的合成有关。几分钟后,早期基因的转录变慢,晚期基因开始表达,合成基因编码病毒的结构蛋白及与装配等有关的非结构蛋白。感染后约6分钟,病毒DNA的复制开始。再过几分钟,复制的DNA分子库和结构蛋白已经积累既,并开始病毒粒结构的装配。把DNA包装入头壳内,就形成完整的头部,与此同时,尾和尾丝的结构也分别产生。这时,已经形成的完整的头部与尾部连接,尾丝则连接到尾板上,于是希活古食计城样烈负就产生了一个感染性噬菌体颗粒。这种颗粒在感染后 12分钟左右即可看到,并继续积蓄25~30分钟。这个周期终结时,合成一种溶菌酶,使细菌裂解,而将成熟的细菌病毒释放。整个过程约需30分钟。
至于RNA和ssDNA的细菌病毒,各有其独特的复制过程,线状噬菌体增殖的子代可不裂解宿主细菌而游离出来。
噬菌体DNA注入宿主细菌后京达财茶,即进入营养状态,完成细菌病毒的增殖周期,这类细菌病毒称为烈性噬菌体。还有扩方延应结愿跑一类细菌病毒,在感染细菌后也可不选择上述增殖的途刑东还什略科直鲜顶笑谓径,而以其DNA整合到细菌染色体中,成为前噬菌体状态。这时也不能用任何方法在细菌内检出细菌病毒颗粒的存在,细菌继续生存并进行分裂繁殖,前噬菌体则随着细菌染色体复制而传递给它的后代。如此衍生的菌株的每个细菌染色体都带有这种前噬菌体结构,称为溶原性菌,这类细菌病毒则称为温和性噬菌体。因此,温和性噬菌体感染后可出现两种反应,即裂解性反应和溶原性反应。温和性噬菌体形成的噬斑总是长有一薄层菌而呈半透明状,这是由于在被感染的细菌中,总有一部分被裂解,另一部分被溶原化,而溶原化的细菌会存活下来并继续生长。温和性噬菌体种类极多,同一菌株也可由多种不同的噬菌体溶原化。温和性噬菌体与烈性噬菌体在遗传上是不同的,温和性噬菌体具有能编码合成一种称为阻遏体蛋白的基因,这种阻遏体蛋白能阻止细菌病毒所有有关增殖基因的表达,使其不能进入上述的营养状态;前噬菌体在细菌的染色体上不能增殖,细菌也不发生裂解,而成为溶原性细菌。另外还有一些基因调节控制阻遏性的合成,以维持稳定的溶原状态。如果阻遏体活性的水平降低到不足以维持溶原状态的程度,前噬菌体又可离开染色体进入增殖周期,这个细菌又可产生细菌病毒。这种现象在溶原菌培养物中是以较低的频率自发产生的,如果加以人工诱导,可使大多数溶原细菌产生细菌病毒而裂解。起诱导作用的有紫外线和多种射线以及致突变、致畸、致癌或抗癌的化学药物。阻遏体蛋白除阻遏前噬菌体的基因组外,同样能阻遏进入溶原性菌的其他同型细菌病毒的基因组,使其不能在该菌内增殖。因此,溶原性菌对同型细菌病毒呈现一种特异的免疫现象。
溶原性菌除具有产生细菌病毒的潜力和对相关细菌病毒的免疫性之外,有时还伴有其他性状的改变。称为溶原性转换。现在知道很多菌类的不同性状受到溶原性的影响。例如,白喉杆菌产生毒素是由于其前噬菌体带有毒素蛋白的结构基因;金黄色葡萄球菌某些溶血素的产生与溶原性有关;沙门氏杆菌、痢疾杆菌等抗原结构也与溶原性有关。
由溶原性细菌染色体上切离的前噬菌体,如果在细胞内未能进入营养状态,从而本身未能增殖,宿主细菌仍继续生存和繁殖,这样衍生的菌株就失去了前噬菌体和溶原性,称为复疗菌株。因溶原化而改变的某些性状也会随着溶原性复疗而恢复。
当溶原性菌中的前噬菌体因某种已知(如诱导)或未知(自发的)原因离开宿主染色体并进行增殖时,有少数细菌病毒基因组将邻近宿主的基因带下(而噬菌体DNA的另一端则失去一个片段),当其感染另一菌株时,可将宿主基因导入并发生重组而表现出这个基因的性状的现象。
细菌病毒因其所具有的特性而成为探讨核酸(DNA和RNA)的复制、转录、重组,基因表达的调节控制,病毒与宿主的关系等各方面的研究对象,促进了病毒学、分子生物学、遗传学的发展。作为基因的载体,在基因工程的研究中也成为有用的工具。细菌病毒依靠特异性的裂解作用,可极其精细地将致病菌分为不同型,这对细菌性疾病流行病学的传染源追踪是极为有用的。在临床医学中,曾试用细菌病毒诊治某些细菌性感染的疾病。可利用对溶性菌的诱导现象进行抗肿瘤药物的筛选和致癌物质的检查。细菌病毒的污染可能给发酵工业(如食品工业、抗生素工业等)造成很大损失,因此,防治这种污染也是一项重要的工作。
一些细菌性病毒例如绿脓杆菌噬菌体,可以治疗相关疾病。烧伤病人容易感染绿脓杆菌,引起化脓性感染,不容易控制,科学家利用绿脓杆菌噬菌体却可以有效治疗烧伤病人的化脓性感。因为绿脓杆菌噬菌体是细菌性病毒,在绿脓杆菌的体内生活繁殖,使绿脓杆菌无法生活。
 细菌病毒
细菌病毒
 关注微信
关注微信