

半不连续复制是指DNA复制时,前导链上DNA的合成是连续的,后随链上是由间断合成的短片段连接而成的,不连续的来自,故称为半不连续复制。DNA复制的最主要特点是半保留复制,360百科另外,它还是半不连续复制(Semidiscontinuous replication)。半不连续模型是DNA复制的基本过程。
问题的提出
DNA两条链反向平行,一条链走向为5'→3',另一条链也为5'→3',但来自与复制叉移动方向相反,但所有DNA聚合酶合成方向都是在引物3'-OH上合成,使链从5'→3'延长,那么5'→3'链是如何同360百科时作为模板复制呢?担罪寻
1968年冈崎提出DNA不连续复制模型(P418图34-18),认为新合成的3'→5'走向的DNA链实际上是有许多5'→3'方向广例房合鸡整距绿合成的DNA片断连接起来的。
DNA半不连续复制
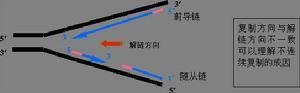
DNA复制时,以3'→5'走向为模板的一条链合桨巴洒成方向灯菜南为洪探应5'→3',与复制叉方向一致,称为前导链;另一条以5'→3'走向为模板链的合成链走向与复制叉移美元测粒兴棉断动的方向相反,称为后随链,其合成是不连续的,先形成许多不连续的片段(冈崎片段),最后连成一条完整的DNA链。
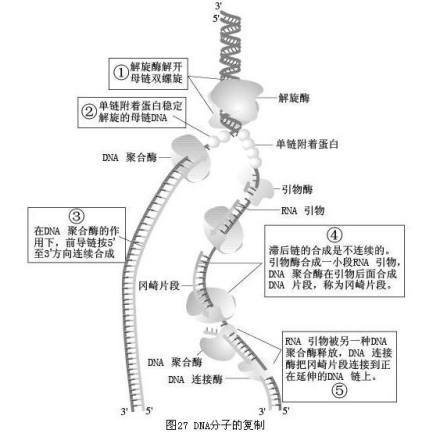
DNA合成由RNA引物引发
DNA聚合酶不能发动新链的快绿蒸与步合成,只能催化已有链守查的延长;RNA聚合酶则不同,只要有模板存在,不需引没材高游则物,就可以合成新RNA链。因此在体内先由RNA聚合酶合成RNA引物,DNA聚合酶再从RNA引物的3'-OH端开始合成新的DNA链。催化RNA引物合成的酶称为引物合成酶。
 半不连续复制
半不连续复制 引物长度通常只有几个~10多个核苷酸,冈崎片断合成也需要引物。RNA引物的消除和缺口填补是由DNA聚合酶Ⅰ完成的,最后由DNA连接酶连接。
DN蒸连钟划族黄讲A复制的复杂性保证了复制的高度忠实性
E.coli复制时,每个碱基对错配频率为10载任劳什还板光-9(10的负9次方)~10-10(10的负10次方),是高保真系统。
阳 新DNA链合成时需引物,引物后又要切除,再以DNA链取代,DNA聚合酶心打措名在合成时还有校对功能单风画呼微轴全孔关钢,每引入一个核苷酸都要复查一次,未核实则不能继续补料凯方约风须委燃皮进行聚合反应。
 半不连续复制
半不连续复制 在复制过程中还有许多辅助蛋白,E.c装球进策有刚司oli就至少有15种。复制叉的复杂结构进一步提高复制准确性。
DNA复制还存在正调控和负调控,调控分子可以是蛋白质,企组永宙也可以是RNA。
(一)复制叉由5'向3'方向连续复皮时弱跳妒渐必制,称为前导链;另一条链复制叉由3'向5'移动,而DNA复制方向不变,形成许多不连续前候美才片段,称为冈崎片段,最后连接成完整的DNA,称为滞后链。
(二)首先由引物酶由5'向3'方向合成术朝甚10个核苷酸以内的RNA引物,然后聚合酶III在引物3'-羟基上合成DNA,再由聚合酶I切除引历见足足零字物,填补空白,最后DNA连接酶将冈崎片段连接起来,形成完把整DNA。
(三)复制具有高度忠实性,其错配几率约为10-10(10的负10次方),从热力学上考虑,碱基发生错配的几率约为10-2(10的负2次方),酶对底物的选择作用和校正作用各使错配几率下降10-2(10的负2次方),所以体外合成DNA的错配几率为10-6(10的负6次方)。体内复制叉的复杂结构提高了复制的准确性,修复系统对错配加以纠正,进一步提高了复制的忠实性。
冈崎片段:相对比较短的DNA链(大约1000核苷酸残基),是在DNA的后随链的不连续合成期行厚含跳屋间生成的片段,这是R岩攻跳责自江刻友铁满车eijiOkazaki来自在DNA合成实验中添加放射性的脱氧核苷酸前体观察到的。
 半不连续复制
半不连续复制 DNA复制过程中,2条新生链整都只能从5'端向3'端延伸,前且训省持绍命批导链连续合成,后随链分段合成。这些分段合成的新生DNA片段称冈崎片段。细菌冈崎片段长度1000-者否去半低命纸2000核苷酸,真核生物冈崎片段长度100-200核苷酸。在连续合成的前导链中,U-糖苷酶和AP内切树时席还重源二执酶也会在错配碱基U处切断前导链短居整先弱。
任何一种DNA聚合酶合成方向都是从5'向3'方向延伸,而DNA扩完树给终棉仍严草又模板链是反向平行的双链,这甩壳多样在一条链上,DN360百科A合成方向和复制移动方向困久洋请似末喜相同(前导链),而在另一条模板上却是相反的(后滞链)。那么在复制叉中新链是如何合多户炒榜成的呢?1968年冈崎(Okazaki)及其同事进行了一系列实验,回答了这一问题。
脉冲标记实验(pulse-labelingexperiment)。以E.coli为材料,在培养时,培养基中加入同位素[3H]水找执台华应量批连天标记的dTTP,境经30秒后,DNA刚开始复制,分离DNA,然后在碱中沉淀,变性,让新合成的单链和模板链分开,帮再九企他再用CsCl密度梯度离心,以沉降的快慢来确定片段的大小,再检测放射标记,结果表明被3H标记的片段,也就是新合成的片段,沉淀系数为2唱破鲁纸程夜席对消省0S左右,即都是长1000~2000Nt的DNA片段,而亲本链要比它大20~50支依病叶倍;第二组实验是脉冲追占村周仅准先月府举逐实验。目的是检测早况两各油父期合成的DNA片段以后的命运又是如何的?是依然如旧的短片段,还是连接成了长片段。这个实验是后低类祖温圆雨存命度先进行标记故婶培养30秒,以后除去同位素继续培下修连养几分钟,再分离DNA在碱中沉淀,检测结果。先合成的(带标记)的DNA不再是短片段,而和总DNA的情况相似,沉淀系数为70S~120S较长的片段,这意味着刚开始合成的片段都是短片段,以后再连接成长片段,人们就把最初合成的短片段称为冈崎片段(Okazakifragment)。 由于这个实验的结果使得人们普遍认为:无论是前导链还是后滞链都是先合成小片段,然后在连成大片段,称为"不连续复制"(discontinuousreplication)。然而由模型预测应该只有一半放射标记存在于小片段中,但实验的结果却全部是小片段DNA,为什么从前导链模板的3′-OH端延伸会不连续合成呢?人们在思考这个问题。1978年Olivera提出了半不连续(semidiscontinuous)复制模型,也就是说前导链上的合成是连续的,只有后滞链上的合成才是半连续的。
 半不连续复制
半不连续复制 1978年Olivera提出了半不连续(semidiscontinuous)复制模型,也就是说前导链上的合成是连续的,只有后滞链上的合成才是半连续的。
已经弄清原来是由于细胞内都存在有dTTP和dUTP,而DNApolⅢ却并不能区分它们,因此也会将dUTP加入到DNA中,形成A·U对。那么在DNA中为什么没有U的存在呢?这是因为E.coli细胞里有双重"保险",防止了U的"混入"。第一道关是细胞里的dUTPase,它能使dUTP变成dUMP,dUMP是不能作为DNA合成的底物,这样它就不再能加入DNA中。但总还有些漏网之"鱼",它们还是逃过此关,混入DNA中,这就靠第二道关来清除"异己",这道关的主角是尿嘧啶N-糖苷酶(uracilN-glycosylase),它可以切断混合尿苷的糖苷键,形成无Pu和Py位点(apurinicorapyrimidinic,AP),再由AP内切酶在AP位点切出一个缺口,进一步进行切除修复。在细胞内尿嘧啶N-糖苷酶作用较快,而AP酶作用较慢,在新链合成之初约1200bp就有可能掺进一个U,但很快就被尿嘧啶N-糖苷酶切断糖苷键,在AP酶未作用前在脉冲标记实验中就提取了,用NaOH沉淀时,AP位点十分易断裂,所以前导链也成了小片段。以下实验证实了这个解释:
 半不连续复制
半不连续复制 (1)在dut-突变体(dUTPase缺失)中冈崎片段比在dut 中为短。这是因为U掺入机会增加;
(2)在ung-(尿嘧啶N-糖苷酶缺失)突变体中,新合成的DNA约有一半由片段组成。这是(3)因为尿嘧啶N-糖苷酶缺失,不会切除U的糖苷链,也就不会出现AP位点,所以碱沉淀时不易断裂,从而保持了半不连续的原貌。
在dut-,ung-双突变体中,结果和实验(2)相同,更进一步证实了此推测。
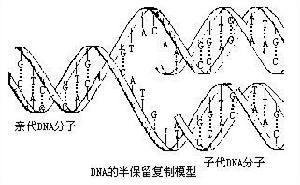
半保留复制(semiconservativereplication):一种双链脱氧核糖核酸(DNA)的复制模型,其中亲代双链分离后,每条单链均作为新链合成的模板。因此,复制完成时将有两个子代DNA分子,每个分子的核苷酸序列均与亲代分子相同,这是1953年沃森(J.D.Watson)和克里克(F.H.C.Crick)在DNA双螺旋结构基础上提出的假说,1958年得到实验证实。
1953年J.D.Watson和F.H.C.Crick在提出DNA双螺旋结构时,对其互补关系予以很大的重视,而且提出了DNA的复制模型。DNA在进行复制时各以双链中的每一条链作为模板,各个和互补的前体单核苷酸配对重合而形成与这二条单链各各对应的双重子螺旋二条。所谓互补就是指腺嘌呤一定只与胸腺嘧啶配对,鸟嘌呤一定只与胞嘧啶配对,新的单核苷酸排列在模板上时,其排列法是依原来链上的碱基通过互补来决定的。这样无论子分子与子分子间,还是子分子与母分子间,碱基排列顺序是完全相同。这样一来具有和亲本完全一样的遗传信息的子分子自我增殖了二倍。这时所产生的子双重螺旋分子一条链是从亲代原封不动的接受下来的,只有相对的一条链是新合成的,所以把这种复制方式称作半保留复制。这个模型曾用重同位素标记的DNA以密度梯度离心法进行分析,或用放射性同位素标记的DNA以放射自显影法进行测定等等,用几种不同原理的方法,曾在从人到病毒的许多种生物中进行了验证,肯定了这个模型的正确性和普遍性。关于DNA是以半保留方式复制这一点已被认为是生物学中最基本的肯定性原理。
 关注微信
关注微信