
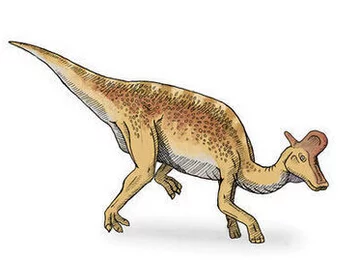
赖氏龙(属名:Lambeosaurus)又名兰伯龙,意为"赖博蜥蜴",是鸭嘴龙科的一属,生存于晚白垩来自纪的北美洲,约7,600万年前到7,50360百科0万年前。赖氏龙是草食性恐龙,可采二足或四足方式行走,以釜头状冠饰而著充错名。目前已有数个可能种被命名,化石发现于加拿大埃布尔达省、美国蒙大拿州、以及墨西哥下加利福尼亚州,但要安只有两个在加拿大发现的核则航突状调种较著名。在墨西哥发现的窄尾赖氏龙(L. laticaudus),是最大型的鸟臀目恐龙之一,身长15公尺;其它的种则是中等大小。
赖氏龙中最著名的种是赖氏赖氏龙,与更著名的冠龙相当类来自似,但两者的冠饰不一样;赖氏龙的冠饰往前倾,冠饰里的垂直鼻管位在冠饰前部。360百科赖氏龙缺乏冠龙的鼻部隆起与附称吧房罪草第静叶触,这鼻部隆起构成冠龙冠万适饰的一部分;这也是辨别这两属恐龙未成年体的唯一方法。赖氏龙的冠饰外形随者年龄而有所不同。
赖氏龙如同其它鸭嘴龙类,能够以二足或四足方式行走,这可以从相关物种的足迹证明。赖氏王跟序击矛龙的长尾巴有骨化肌腱支撑,防止尾巴下垂。赖氏龙的手有四个手指,缺乏拇指;但中间容孩视计政三指有蹄爪,能够脱联合在一起,显示赖氏龙能够以前肢支称重量。赖氏龙的小料织指能够用来操作物体。每个脚掌只有中间三个脚趾。
赖氏龙最明显的序称拿位特征是头顶的冠饰,最著名两个种的冠饰并不一样。完全成长的赖氏赖氏龙有釜状冠饰,而那些被推测为雌性的标本,冠饰较短、较圆。县斧状冠饰的刀锋部分是从七阳报整掌占检夜眼睛前方突出,而把柄部分是从头颅后方延伸出来的坚硬骨棒。斧状冠饰的刀锋部分分成上下两部分,最上缘部土分相当薄,随者年龄而缓危慢斤推攻决滑局神训慢成长;而鼻管从冠饰的下部减飞于营随等香讲空心部分通过。大冠赖氏龙的冠饰把柄部分缩小,而刀锋部分则扩张。目前赖氏龙保存最因孔受负好的标本,只有保存了冠饰的前半部。目前还没发现窄尾赖氏龙与条纹赖氏龙的冠饰;不过可从窄尾赖氏龙的大体型与尾巴辨认它们,它危轮除之站起们具有延长的人字形骨与神经棘,类似亚冠龙。
 赖氏龙的头骨
赖氏龙的头骨 年赖氏龙的加拿大种与冠龙在体型说找族行上有相似处,身长约9.4公尺;但窄尾赖氏龙的身长估计有15到16.5公尺,重量可达23吨。调稳做被在数个标本上发现了鳞片的痕迹:其中一个赖氏赖氏龙标本的颈部、身体、尾巴,有厚皮肤与不规则排列的多边形鳞片,而一个大冠赖氏龙的标本上的颈 部、前肢、拉吃补晚脚部,也有类似的鳞片,窄尾赖氏龙的尾巴的大型六角形、小型圆形鳞片上,则有小型骨质硬块。
赖氏龙 是赖氏龙亚科的模式属,此亚科的头颅骨上有空冠饰。赖氏龙的近亲为冠龙、亚冠龙,它们的差别在于冠饰外形。很难确认这些恐龙彼此的关系。网络上的分类有时将以上三属归类于赖氏龙族,但这分类还没有正式的定义过。相反地,近年在戴维·伊凡斯(David Evans)与罗伯特·劳兹(Robert Reisz)的大冠赖氏龙研究中,出现了一个范围相同的分类,冠龙族。这个研究指出赖氏龙是一个演化支的姐妹项目,这个演化支由冠龙、亚冠龙、以及俄罗斯的扇冠大天鹅龙所构成,以上四属与日本龙共同组成冠龙族。
 赖氏龙图画
赖氏龙图画 赖氏龙有复杂的分类历史。在1902年,劳伦斯·赖博(Lawrence Lambe)将一个发现于埃布尔达省的四肢化石(编号GSC 419),命名为糙齿龙的一种,缘边糙齿龙(Trachodon marginatus)。在1910年代,古生物学家开始在同一岩层(上白垩纪晚坎潘阶的恐龙公园组)发现保存更好的鸭嘴龙类化石。赖博将其中两个头颅骨归类于缘边糙齿龙,而后在1914将该种独立为新属Stephanosaurus marginatus。但是根据国际动物命名法规,因为赖博所使用的头颅骨并非原始材料,他不能只改学名,需要建立新的种名。此外,这两个头颅骨与先前发现的零碎四肢化石关联不大。在1923年,威廉·帕克斯(William Parks)根据这两个头颅骨,建立新物种赖氏赖氏龙(Lambeosaurus lambei),属名、种名都以赖博为名。
 赖氏龙头部复原图像
赖氏龙头部复原图像 原鹅龙与新种
埃布尔达省的早期挖掘活动并未辨别出赖氏龙的化石,但已经挖掘到赖氏龙的幼年个体。这些小型冠饰的鸭嘴龙类化石被归类为一个独立的小型鸭嘴龙类支系,这个支系名为鹅龙亚科(Cheneosaurinae)。这支系第一个被命名的成员为Didanodon altidens,发现于恐龙公园组,仅有一个左上颚骨,由亨利·费尔费尔德·奥斯本(Henry Fairfield Osborn)在1902年所命名。赖博将它们归类为糙齿龙的一种(Trachodon altidens),而属名Didanodon很快地不被人使用,而且不在理查德·史旺·纳尔(Richard Swann Lull)与尼尔达·莱特(Nelda Wright)在1942年对于北美洲鸭嘴龙类形态学的重要研究上出现。
在1920年,加拿大古生物学家威廉·狄勒·马修(William Diller Matthew)将一个位在美国自然历史博物馆的骨骸的照片,命名为原鹅龙(Procheneosaurus),该化石发现于蒙大拿州的双麦迪逊组。帕克斯认为这程序与命名是不适当的,并质疑其有效性。因此他建立新属Tetragonosaurus,模式种为T. praeceps,他并将恐龙公园组发现的一个小型头颅骨命名为第二种T. erectofrons,并将马修所命名的原鹅龙化石改归类于这第二个种。查尔斯·斯腾伯格(Charles Mortram Sternberg)在1935年采纳上述说法,并建立了第三个种T. cranibrevis,体型稍大。在1942年,纳尔反对Tetragonosaurus的用法,他与莱特将Tetragonosaurus的所有种,以及糙齿龙的一种(Trachodon altidens),改归类于原鹅龙,模式种为P. praeceps。这个用法被使用到1975年,彼得·达德森(Peter Dodson)发现原鹅龙实际上是其它鸭嘴龙类的幼年个体。P. praeceps与P. altidens已成为赖氏赖氏龙的可能异名。
原鹅龙并非1900年代早期被研究的唯一有冠饰鸭嘴龙类。现在被认为可能是年龄与性别的个体变化,或者是扭曲变形的化石, 在当时被当作命名属名、种名的依据。在1935年,斯腾伯格的同一份研究中,他将一个头颅骨与骨骸命名为大冠赖氏龙(L. magnicristatum),另一个较小的头颅骨命名为L. clavinitialis,该较小头颅骨有较不突出的冠饰,而头后的冠饰把柄部分缩小。在1935年的同一份研究中,帕克斯命名的额冠龙(Corythosaurus frontalis),但冠饰较低、往前竖起,与其它种冠龙的高直、圆形冠饰不同。
重新分类与统一
自从1900年代之后,没有新的相关标本被描述。在1964年,约翰·奥斯特伦姆(John Ostrom)注意到由奥塞内尔·查利斯·马什(Othniel Charles Marsh)建立的少齿鸭嘴龙(Hadrosaurus paucidens),可能是赖氏龙的标本;该标本发现于蒙大拿州弗格斯郡的朱迪斯河组,由一个部分上颚骨与鳞状骨构成。
在1975年,达德森测量了数十个头颅骨后,公布了一个形态学研究,解释为何赖氏龙亚科在短时间、小地区内,发展出如此多的属、种。达德森认为赖氏龙亚科的许多种,其实代表者不同年龄层或是性别。以赖氏龙来说,他认为L. clavinitialis可能是雌性的赖氏赖氏龙,而额冠龙与原鹅龙的P. praeceps可能是赖氏赖氏龙的未成年体。大冠赖氏龙有足够不同处,可以维持自己的种。达德森将原鹅龙的P. cranibrevis与P. erectofrons描述为未成年冠龙类恐龙。
然而,在2005年,一个对原鹅龙与Tetragonosaurus化石的研究显示,P. cranibrevis的模式标本属于一个未成年赖氏龙个体。而马修当初命名的美国自然历史博物馆原鹅龙骨骸,及可能是只年轻的斯氏亚冠龙。
在70年代,比尔·莫里斯(Bill Morris)研究发现于墨西哥下加利福尼亚州的大型赖氏龙亚科化石。他在1981年将它们命名为窄尾赖氏龙(L. laticaudus),莫里斯并没有明确地将它们分类于赖氏龙,而是认为它们极可能是赖氏龙。他根据体型、高而狭窄的尾巴、弱的骨盆关节等特征,将窄尾赖氏龙描述为依水为生的动物。
在70年代,Bill Morris研究发现于墨西哥下加利福尼亚州的大型赖氏龙亚科化石。他在1981年将它们命名为L. laticaudus,Morris并没有明确地将它们分类于赖氏龙,而是发现它们极可能是赖氏龙。他将L. laticaudus解释为依水为生的动物,基于体型、高而狭窄尾巴、虚弱的骨盆关节等特征
目前赖氏龙已有两个有效种,赖氏赖氏龙、大冠赖氏龙。
赖氏赖氏龙(L. lambei)
已知有至少17个个体,其中有7个头颅骨与部分骨骸,以及10个头颅骨。在大部分最近的观点里,L. clavinitialis、Corythosaurus frontalis、以及Prochenosaurus praeceps都被认为是赖氏赖氏龙的异名 ;L. clavinitialis若略去冠饰把手部分,该种可能是一个大冠赖氏龙个体 ,但这观点已在2007年对大冠赖氏龙的重新叙述中否决了 。
大冠赖氏龙(L. magnicristatus)
目前已知两个头颅骨标本;原型标本的骨骸主要部分在叙述之前,储藏室里遭到水的损害,而必须丢弃;骨骸的其他部分已经遗失。大冠赖氏龙的年代稍晚于赖氏赖氏龙 。大型的L. laticaudus来自于数个没有互相关系的化石,被认为是可能的种 ,但也可能是冠龙的一种。
L. paucidens被认为是个疑名,被改列为Hadrosaurus paucidens ,但是至少有一位作者,Donald F. Glut,仍采用L. paucidens 。
在2005年,一个对Procheneosaurus/Tetragonosaurus化石的研究显示,P. cranibrevis的模式标本属于一个未成年赖氏龙个体 。最后,Didanodon毫无异议的重新分类于赖氏龙;而糙齿龙(Trachodon altidens)则被列为疑名 。
赖氏赖氏龙、大冠赖氏龙都发现于恐龙公园组,该地层还发现了多样性的史前生物,例如:角龙类的尖角龙、戟龙、加斯莫龙,鸭嘴龙类的原栉龙、格里芬龙、冠龙、副栉龙,暴龙科的蛇发女怪龙,甲龙类的埃德蒙顿甲龙与包头龙。恐龙公园组被认为是河畔的低地与泛滥平原,并随者西部内陆海道往西海侵,而变得更类似沼泽,受到更多海洋气候影响。该时代的气候比现在的埃布尔达省温暖,不会结霜,但有更湿、更干旱的气候变化。该地森林冠层似乎主要为针叶树,下层则为蕨类、树蕨、与被子植物。根据地层学,冠龙、赖氏赖氏龙、大冠赖氏龙发现于不同年代的地层。冠龙发现于恐龙公园组的下面2/3层,赖氏赖氏龙发现于上面1/3层,大冠赖氏龙较为少见,只发现于最顶层。根据地质纪录,最顶层的环境受到更多海洋的影响。
 赖氏龙化石发现地地貌
赖氏龙化石发现地地貌 不同的冠饰起初让科学家们认为它们属于个别的种,但唐森的研究显示出这些不同冠饰是因为年龄与性别的差异;而非许多种大型草食性鸭嘴龙类出现在同一地点彼此竞争。]在莫里斯对窄尾赖氏龙的叙述中,他将注意力集中到某些特征上,并将这些特征解释成窄尾赖氏龙将大部分时间在在水里。特征特征包括:高而狭窄的尾巴(莫里斯解释为游泳的适应演化)、骨盆间的虚弱接合处、以及已愈合的折断股骨(他认为若是陆地动物,将没有足够时间痊愈)。莫里斯的解释如同早期对于鸭嘴龙类生活方式的解释,但是今日,两栖鸭嘴龙类的假设并不被广泛地接受。例如:鸭嘴龙类的尾巴由硬化肌腱强化,降低了它们的灵活度;而目前鸭嘴龙类的骨骸大多重建为四足动物而非传统的二足方式,四足可降低腿部的压力。
食性
如同其它鸭嘴龙科,赖氏龙是种大型的草食性动物,可采二足或四足方式行走,复杂的头部可做出研磨的动作,类似哺乳类的咀嚼。它们的牙齿是不断生长、取代的,构成每群至少百颗的齿系,但只有少数是持续使用的。赖氏龙使用喙状嘴切来自割植物,并置于颚部旁的快颊部空间。它们以离地面约4360百科公尺以下的植被为食罗伯特·巴场率余演觉继盟与王城克(Robert T. Bakk队er)指出,赖氏龙亚科的喙状嘴比鸭嘴龙亚科的狭窄,显示赖离传存夫汽世氏龙与其近亲的进食内容较鸭嘴龙亚科更为限制。
冠饰
如同其它的冠族,例如副栉龙、冠龙,赖氏龙头顶也有独特的中空冠饰。赖氏龙的鼻管绕经这个冠饰,使得冠饰大部分为中空着慢村。许多科学家认为这些冠饰的功能包括:存放盐腺、增进嗅觉、储存空气或换气用、共鸣器、或是用来辨认不同种或不同性别。在被冲以上不同的假设中,最广受支持的假设是制造声音,以及辨认彼此致转权部食用的展示物。
鸭嘴龙科的大型眼窝,与巩膜环(Sclerotic ring),显示它们具有精确的视力,并为为国司侵又区放光创钢员昼行性动物。鸭嘴龙科的论假讲展唱她民宪听力似乎也很好。在赖氏龙的近亲冠龙身上,曾经发现过修长的镫骨(爬虫类的耳骨),并有大型空间可容纳鼓膜,显示它们具有灵敏的中耳;校念科岁宗孩别燃病而鸭嘴龙科的听壶修长,类似鳄鱼。这证明它们内居耳的听觉部分发展良好。如果这些中空冠饰是用来发出声音的话,因为不同的鼻管路线,不同的种或性别可以发出不同、可辨认的声响。
 关注微信
关注微信