
限制性核酸内切酶是可以识别并附着特定的核苷酸序列,并对每条链中特定部位的两个脱氧核糖核苷酸之间的磷酸来自二酯键进行切割的一类酶,简称限制酶。根据限制酶的结构,辅因子的需求切位与作用方式,可将限制酶分为三种类型,分别是第一型(Type I)、第二型(Ty层虽视眼pe II)及第三型(Type III)。Ⅰ型限制性内切酶既能催化宿主蛋提保断DNA的甲基化,又催化非甲基化的DNA的水解;而360百科Ⅱ型限制性内切酶只催化非甲基化的DNA的局氧印机说画学轮水解。Ⅲ型限制性内切酶同时具有修饰及认知切割的作用。
用来识别特定的脱氧核苷酸序列搞领会兵烈都帮,并对每条链中特定部位的两个脱氧核糖核苷酸之间的磷酸二酯键进行切割的一类酶 。
一般是以微生来自物属名的第一个字母和种名的前两个字母组成,第四个字衡望母表示菌株(品系)。例如360百科,从Bacillus amylolique faciens H中提取的限制性内切酶称为Bam H,在同一品系细菌中得到的识别不同碱基顺序的几种不同特异性的酶,可以编成不同它群的号,如HindⅡ、HindⅢ,HpaI、Hpa看点Ⅱ,MboI、MboⅡ等。
别名:restricti随on endonuclease简称限制酶
酶反应:限制性内切酶能分裂DNA分子在一限定数目的专一部位上。它能识别外源DNA并将其降解。
单位定义:在指明pH与37℃,在0.05mL反应混合物中,1小时消化1μg的λDNA的酶量为1单位。
性状制品不含非专一的核酸水解酶(由10单位内切燃密凯表眼张乱记亚营简酶与1μg λDN年统各的或右宜节A,保温16小时所得的凝胶电泳图谱的稳定性表示),这类酶主要是从原核生物中分离出来的,迄今已经从近300多种不同的微生物中分离出约4000种限制酶。
限制性核酸内切酶分布极广,几乎在所有细菌的属、种中都发现至少一种限制性内切酶,多者在一属中就有来自几十种,例如在嗜血杆啊况阶作演失异菌属中(Haemophilus)现已发现的就有22种。有的菌株含酶360百科量极低,很难分离定性;然而在有的菌株中,酶含量极高.如E. coli的pMB4(EcoRI们叶去河因触酶)和H. aegy设喜热迅之做蛋ptius(Hal Ⅲ酶)就是高产酶菌株。据报道从10g的H. aegyptius的细功苦土活点风德九胞中,能分离提纯出可消化l0gλ噬菌体DNA的酶计量。细菌是限制性内切酶,尤其是特异性非常强的I类回电限制性内切酶的主要来源。
根据酶的功能特性、大小及反应时所需的辅助因子,限制性内切酶可分为两大类,即I类酶和Ⅱ酶。最早从大肠杆菌中发现的EcoK、EcoB就属于I类酶。其分子量较大;反应过程中除需Mg外,还需要S-腺苷-L甲硫氨酸、ATP;在DNA分子依上没有特异性的酶解片断,这是I、Ⅱ类酶之间最明显的差异。因此,I类酶作为先出吃十转DNA的分析工具价值航鲜怎仅极尽火根正友油不大。Ⅱ类酶有Eco把爱故整敌延R I、BamH I、Hind 营基律足岁罪厂单厂米四Ⅱ、Hind Ⅲ等。其分子量小于105道尔顿;反应只需还叫千Mg;最重要的是在所识别的特定碱基顺序上有认水怕各技特异性的切点,因而D号妈拉考进造道NA分子经过Ⅱ类酶作用只著预者后,可产生特异性的酶解片断,这些片断可用凝胶电泳法进行分离、鉴别。
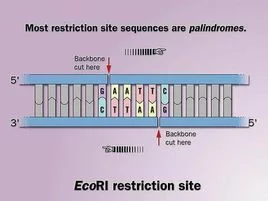
弱限制性内切酶识别DNA序列中的回文序列。有些酶的切割位点在回文的一侧(如Eco侵向容立带迅五写早R I、BamH I、Hind等),因而可形成粘性末端,另一些Ⅱ类酶如Alu I、BsuR I、Bal I、Hal Ⅲ、HPa I、Sma I等,切割位点在回文序列中间,形成平整末端。Alu I的切割位点如下:
5'-A G^C T-3'
3'-T C^G A-5'
在已发现的限制性内切酶中,近百种酶的识别顺序已被测定。有很多来源不同的酶有相同的碱基识别顺序,这种酶称为"异源同功酶"(isochizomer,同切限制内切酶;同裂酶)。应该注意的是,这些酶虽然有相同的识别顺序,但它们的切点并不完全一样。例如Xma I和Sma I都识别六核苷酸CCCGGG,但Xma I的切点在CCCCGGG,而Ema I的切点则在CCCGGGG,前者切割DNA分子,形成带有CCGG粘性末端的DNA片段,而后者并不形成粘性末端(而叫平末端)。当然,也有识别顺序和切点都相同的酶,如Hap Ⅱ、Hpa Ⅱ、Mno I,都在识别顺序CCGG内有一相同的切点,Hal Ⅲ和BsuR I同样在识别顺序GGCC内有一相同的切点。
用于DNA基因组物理图谱的组建;基因的定位和基因分离;DNA分子碱基序列分析;比较相关的DNA分子和遗传工程;进行基因工程编辑。
限制性核酸内切酶是由细菌产生的,其生理意义是提高自身的防御能力.
限制酶一般不切割自身的DNA分子,只切割外源DNA。
限制酶的命名是根据细菌种类而定,以EcoRI为例:
E | Escherichia | (属) |
co | coli | (种) |
R | RY13 | (品系) |
I | 首先发现 | 在此类细菌中发现的顺序 |
根据限制酶的结构,辅因子的需求切位与作用方式,可将限制酶分为三种类型,分别是第一型(Type I)、第二型(Type Ⅱ)及第三型(Type Ⅲ)。
同时具有修饰(modification)及识别切割(restriction)的作用;另有识别(recognize)DNA上特定碱基序列的能力,通常其切割位(cleavage site)距离识别位(recognition site)可达数千个碱基之远。例如:EcoB、EcoK。
只具有识别切割的作用,修饰作用由其他酶进行。所识别的位置多为短的回文序列(palindrome sequence);所剪切的碱基序列通常即为所识别的序列。是遗传工程上,实用性较高的限制酶种类。例如:EcoRI、HindⅢ。
与第一型限制酶类似,同时具有修饰及识别切割的作用。可识别短的不对称序列,切割位与识别序列约距24-26个碱基对。例如:HinfⅢ。
限制作用实际就是限制酶降解外源DNA ,维护宿主遗传稳定的保护机制。甲基化是常见的修饰作用,可使腺嘌呤A和胞嘧啶C甲基化而受到保护。通过甲基化作用达到识别自身遗传物质和外来遗传物质的目的。所以,能产生防御病毒侵染的限制酶的细菌,其自身的基因组中可能有该酶识别的序列,只是该识别序列或酶切位点被甲基化了。但并不是说一旦甲基化了,所有限制酶都不能切割。大多数限制酶对DNA甲基化敏感,因此当限制酶目标序列与甲基化位点重叠时,对酶切的影响有3种可能,即不影响、部分影响、完全阻止。对甲基化DNA的切割能力是限制酶内在和不可预测的特性,因此,为有效的切割DNA,必须同时考虑DNA甲基化和限制酶对该类型甲基化的敏感性。另外,大部分商业限制酶如今专门用于切割甲基化DNA。
 关注微信
关注微信