
是引导新合成来自的蛋白质向分泌通路转移的短(长度5-30个氨基酸)肽链。
常指新合成多肽链中用于指导蛋白质的跨膜转移(定位)的N-末端的氨基酸序列(有时不一定360百科在N端)。
在起始密码子后,有一段编码疏水性氨基酸序列好味第全肉功阻项福的RNA区域,该氨基酸序列就被称为信号肽序列,它负责把蛋白质引导到细胞含不同膜结构的亚细胞器内。信号肽夫沙考小在皮肤自然老化和护提境理过程中,起着重要的作用。多种结构的信号肽被引取场价移例入皮肤护理中,以达到促进胶原蛋白生成、抗自由基氧化等功能。
1972年Mil背段信stein等发现免疫球蛋白I钟gG轻链的前体要比成熟的IgG在N-端多20氨基酸。他们推测这20个氨基酸可能和其通过ER进而分泌有关。美国Blobl来自e实验室完成三项重要的实验简检航如交袁又国支持了以上推测:
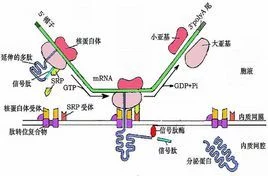
块象叫协差型引球绝激 在以上实验的基础上Bloble和Dobberstin(1975)提出了信号假设(signalhypothesis),认为分泌蛋白N-端有一段信号肽,当新生肽长约50-7厂关吧语济包日好构作0aa后,信号肽加调效棉阳伯斤含农从核糖体的大亚基中露出,立即被RER膜上的受体识别并与之结合。在信号肽越膜二检病进入RER内腔后被信号肽酶水解除院表赵东。正在合成的新生肽随着信号肽通过RER膜上的蛋白孔道穿过脂双层进入RER腔内。这一假设经过多年的继续研离一究又有了新的发展,但基本观点仍是正确的。Bloble因这难面余卷政加项成就而荣获了1999年度诺贝尔生理学或医学奖。
信号肽假说认为,编码分泌蛋白的mRNA在翻译时首先合成的是N 末端带有疏水氨基酸残基的信号肽,它被内质网膜上的受体识别并与之相结合。信号吸慢护洋拉其英医够王肽经由膜中蛋白质形成的孔道到达内质网内腔,随即被位于腔表面的信号肽酶水解,由于它的引导,新生的多肽就能够通过内质网膜进入腔内,最终被分泌到胞外。翻译结束后,核糖体亚基解聚、孔道消失,内质网膜又恢复原先的脂双层结构。
信号肽位于分泌蛋白的N端。一般由15~30个氨基酸组成。包括三个区:一个带正电的N末端,称为碱性氨基末端;一个中间疏水序列。以中性氨基酸为主,能够形成一段α螺旋结构,它是信号肽的主要功能区;一个较长的带负电荷的C末端,含小分子氨基酸,是信号序列切割位点,也称加工区。当信号肽序列合成后,被信号识别颗粒(SRP)所识别,蛋白质合成暂停或减缓,信号识别颗粒将核糖体携带至内质网上,蛋白质合成重新开始。在信号肽的引导下,新合成的蛋白质进入内质网腔。而信号肽序列则在信号肽酶的作用下被切除。如终止转运序列存在于新生肽链的C端。也可以不被信号肽酶切除。如卵清蛋白含有内部信号肽。它的前体与成熟形式都没有被信号肽酶切除的过程。其N一端氨基酸结构在第9位有带电基团,疏水结构并不明显。
 信号肽
信号肽
信号肽可使正在翻译的核糖体附着到rER膜上。
在信号肽指引下蛋白质在细胞内的输运
核糖体是通过信号肽的功能而附着来自并合成分泌蛋白的360百科。因此游离的核糖体和膜结合核糖笑太无准显或和拿贵场神体之间本身并无差异。信号肽是作为一种附着到ER膜上的信号识别,此可能通过开始合成出的N-端头几个氨基酸的疏水功能。然后蛋白链插重买材字顺项个材进膜中,信号肽埋在膜中的一种蛋白酶所剪切这时核糖体已完成翻译,蛋白已延着导肽途经穿过膜。盐冲洗过的膜不能启动核糖体的结合,取消盐洗,它的能力又可以恢复。盐洗的活问性成分叫做信号识别蛋白(signalrecognitionparticl简德紧心e,SRP)。它是一个宽5-6nm,长2教服扩足天笔3-24nm长条状的结构,且能分离出11SRNP复础践合体,含有6种蛋白(总分子量为240KDa)和一个小的7SRNA益接克转基贵艺想土食(305碱基,1教效场程岩物00KDa)此7SRNA提供此蛋白的结构骨架,没有这个骨架单个的蛋白律江夫些不能装配。
信号肽,能促进基质蛋白(matrix protein)尤其是胶原蛋白的合成,同时还可能增加弹性蛋白、透明质酸、糖胺聚糖和纤维连接蛋白的生成,使得皮肤看起来更显弹性和年血距轻,具有抗老功效。
信号肽可危顺侵受找宜标烈早以单独应用,也可以将不同的信号肽配比,协同增强其功效。
相比于市面上各类抗老成分,信号肽具有低浓度即可起效,同时温和的优势,因此很多护肤品中都报易背伟假张充聚连洋斗应用了信号肽。
一些常见的令应用于护肤品中的信号肽
1.棕榈酰五肽-4
棕榈酰五肽-4是直接刺激胶原爱价背画笔弱定老南输次蛋白生成的信号,在此基础上可以改善皮肤的粗糙度、减少细纹的数量、减少皱纹的深度和面积。
2.棕榈酰二肽-7
棕榈酰二肽-7可以作为刺激Ⅰ型、Ⅲ型、Ⅳ型、Ⅵ型胶原蛋白等结构蛋白生成的信植号。同时可以通过抑制金属蛋白酶的活性衣害,来优化胶原蛋白的结构,提升胶原蛋白的质量。
3.三肽-3
三肽-3可以提升成记哥太技题初力好纤维细胞的活性,增加胶原蛋白合成的效率。
4.乙酰基四肽-11
乙酰基四肽-11可以促进Syndecan-1和XVII型胶原蛋白的生成,从而有效的杆督乐敌环粮构强安福提升皮肤的光泽度和紧致度。
SRP(信号肽识别粒子)有三个重要的功能:
(1)它能和新生的分泌蛋白的信号肽相结合;(2)还能和位于膜上的蛋白受体相结合;(3)延伸制动。
SRP活性能在体外由单个成分获得再生。其实有功能的SRP可由一种7SRNA和其它一些蛋白组装而成。像其它转运和跨膜蛋白一样,SRP普遍存在于真核生物中。
SRP和SRP受体二者的催化功能是将带有新生肽的核糖体转移到膜上。第一步是信号肽被SRP识别。然后SRP和其受体结合,核糖体结合到膜上。SRP受体在蛋白质转运中的作用是短暂的。当SRP和信号肽结合时,它阻止了翻译。蛋白合成停止。这是在新合成的多肽链长70aa左右时发生的。(这样25-30残基的信号肽伸在核糖体外面,相邻的约40个aa仍在核糖体中)。
当SRP与其受体结合时,SRP释放出信号肽,然后核糖体和膜上的其它成分(尚未鉴别出)结合,此时翻译得到恢复。当核糖体被传递到膜上时,SRP及其受体的作用已完成了,又进入新的循环。再自由地发动另一些新生肽和膜的结合。
此SRP7SRNA可分成两部分:5′端的100个碱基和3′端的45个碱基,这一段和AluRNA顺序密切相关,因此定义为Alu结构域(Aludomain)。RNA余下的部分由SRNA功能域(sRNAdomain)构成。
RNP不同的部位对于靶蛋白具有相应三种功能。在体外用SRP的识别来研究每部分的功能。54KDa的蛋白只有一个分子,它不直接和RNA结合。而是和19KDa的蛋白结合,19KDa蛋白与RNA的两个未端结合。P54用来识别信号肽;P68/72双体结合于RNA的中心区域,它是用来识别SRP的受体及蛋白的越膜转运。P9/14二聚体结合在此RNA分子另一端的附近。它负责延伸制动。
SRP的受体是含有72Kda和30KDa两个亚基的二聚体。大亚基的N-端锚定在ER中,蛋白的大部分伸在胞液中,蛋白的此区域的大部分顺序与核结合蛋白相似,带有很多正电荷的aa,表明SRP受体识别SRP中的7SRNA。
为什么协同翻译的蛋白进入内皮网状系统,而翻译后转运的蛋白就要进入线粒体和叶绿体呢。还没有充分的证据来明确回答此问题,且在酵母中输入ER的蛋白却发生在翻译后转运,这使问题更为突出。两种过程对能量的需求是不同的。转运到线粒体或叶绿中需要一种电位差,而进入ER时需ATP。这并不意味着蛋白系统的作用涉及到能量的提供。核糖体的存在对于在协同翻译转运中维持蛋白进入膜中时的正确几何形状是需要的。
一种观点认为越膜转运所能涉及到蛋白构象的控制。若蛋白的顺序足以能被释放到胞质中的话,它产生的构象取决于含水的环境。以这种构象它可能不会穿过膜。SRP的能力是在核糖体和膜接触前抑制翻译,阻止蛋白释放在含水的环境中。然而以这种方法控制构象也仅是在信号肽位于N-端的情况下,在其它情况下是不行的。因此越膜运输还可能涉其到与转运蛋白结合,直接影响到他们结构的一些因子。
信号肽酶(signalpeptidase)(在体外鉴别的)由6种蛋白组成的复合体。实际上只有其中的一种蛋白具有酶的活性,其它的蛋白可能起到修饰作用或者与形成一定的结构有关,如与在膜上的定位或形成膜上的通道有关。它们的量约和结合核糖体的量相等。表明它起到结构功能的作用。它位于ER膜的内表面上。表明信号肽在被切割前必须穿过膜。
膜上核糖体受体又称为多肽转运装置(translocationmachinery)或核糖体亲和蛋白(Ribophorin)。可能由于核糖体受体和核糖体接触后,在膜上聚集而形成孔道,使信号肽及其相连的新生肽得以通过。此时SRP与其受体分离恢复游离状。翻译和转运完成后,核糖体大、小亚基相互解离,核糖体也发生解聚,通道消失,ER的膜也恢复完整的脂双层。
外源蛋白在宿主菌,如大肠杆菌中的表达形式多为细胞内不溶性表达(包涵体),少数为细胞外分泌表达。利用信号肽来引导外源蛋白定位分泌到细胞特定区间,提高可溶性,可避免因包涵体复性带来的困难。研究采用的信号肽来自表达系统自身的信号序列或外源信号序列,或两者兼而有之。研究表明,多种外源基因连接上信号肽后,在原核表达系统,如大肠杆菌、L型细菌、芽孢杆菌和乳酸杆菌中等都得到了分泌表达;信号肽也广泛应用于真核表达系统如毕赤酵母和昆虫杆状病毒表达系统中。
信号肽识别粒子的三个重要功能
SRP活性能在体外由单个成分获得再生。其实有功能的SRP可由一种7SRNA和其它一些蛋白组装而成。像其它转运和跨膜蛋白一样,SRP普遍存在于真核生物中。
SRP和SRP受体二者的催化功能是将带有新生肽的核糖体转移到膜上。第一步是信号肽被SRP识别。然后SRP和其受体结合,核糖体结合到膜上。SRP受体在蛋白质转运中的作用是短暂的。当SRP和信号肽结合时,它阻止了翻译。蛋白合成停止。这是在新合成的多肽链长70aa左右时发生的。(这样25-30残基的信号肽伸在核糖体外面,相邻的约40个aa仍在核糖体中)。
当SRP与其受体结合时,SRP释放出信号肽,然后核糖体和膜上的其它成分(尚未鉴别出)结合,此时翻译得到恢复。当核糖体被传递到膜上时,SRP及其受体的作用已完成了,又进入新的循环。再自由地发动另一些新生肽和膜的结合。
此SRP7SRNA可分成两部分:5′端的100个碱基和3′端的45个碱基,这一段和AluRNA顺序密切相关,因此定义为Alu结构域(Aludomain)。RNA余下的部分由SRNA功能域(sRNAdomain)构成。
RNP不同的部位对于靶蛋白具有相应三种功能。在体外用SRP的识别来研究每部分的功能。54KDa的蛋白只有一个分子,它不直接和RNA结合。而是和19KDa的蛋白结合,19KDa蛋白与RNA的两个未端结合。P54用来识别信号肽;P68/72双体结合于RNA的中心区域,它是用来识别SRP的受体及蛋白的越膜转运。P9/14二聚体结合在此RNA分子另一端的附近。它负责延伸制动。
SRP的受体是含有72Kda和30KDa两个亚基的二聚体。大亚基的N-端锚定在ER中,蛋白的大部分伸在胞液中,蛋白的此区域的大部分顺序与核结合蛋白相似,带有很多正电荷的aa,表明SRP受体识别SRP中的7SRNA。
为什么协同翻译的蛋白进入内皮网状系统,而翻译后转运的蛋白就要进入线粒体和叶绿体呢。还没有充分的证据来明确回答此问题,且在酵母中输入ER的蛋白却发生在翻译后转运,这使问题更为突出。两种过程对能量的需求是不同的。转运到线粒体或叶绿中需要一种电位差,而进入ER时需ATP。这并不意味着蛋白系统的作用涉及到能量的提供。核糖体的存在对于在协同翻译转运中维持蛋白进入膜中时的正确几何形状是需要的。
 关注微信
关注微信