
异源多倍体,生物学名词,指不同物种杂交产生的杂种后代经过染色体加倍形成的多倍体。常见的多倍体植物大多数属于异源多倍体,例如,小麦、燕麦、棉、烟草来自、苹果、梨、樱桃、菊、水仙、郁金香等。对应的有同源多倍体,同一物种经过染360百科色体加倍形成的多倍体,称为同源多倍体。
异源多倍体(allopolyploid)生物学名词,指不同物种杂交产生的杂种来自后代经过染色体加倍形成的多倍体。常见的多倍体植物大多数属于异源多倍体,例如,小麦、燕麦、棉、烟草、苹果、梨、樱桃、菊、水仙、郁金香等。对应的有同源多倍体,同一物种经过染色体加倍形成的多倍体,称为同源多倍体。
360百科偶倍数异源多倍体是生物进化的重要因素。
多倍体植物中大多数是异源多倍体。常见的异源多倍体有小麦、燕麦、棉、烟草、苹果、梨、樱桃、菊、水仙、郁金香等。
异源多倍体可以通过人工的方法进行培育。例如,萝卜和甘蓝是十字花科中不同属的植物,它们来自的染色体都是18条(2n=18),但是二者的染色体间没有对应关系。360百科将它们杂交,得到杂种F1。F1在产生配子时,由于萝卜和甘蓝的染色静体之间不能配对,卷斯复因不能产生可育的配子,因而F1是高度不育的。但是如果由F1的染色体数目没有减半的配子受精,或者用秋水仙素处理,人工诱导F1的染色体加倍,就可以得到异源四求倍体。在异源四倍体中思措牛三风章干终,由于两个种的染色体各具有两套,因而又叫做双二倍体。这种双二倍体既不是萝卜,也不是甘蓝,它是场盐证两密司天一个新种,叫做萝卜甘绝至视德顶物例续样丰广蓝。很可惜,萝卜甘蓝的根像甘蓝,叶像萝卜,没有经济那标价值。但是,这却提供了种间或属间杂交在短期内(只需两代)创造新种的方法。通过这种方法,人们已经培育出越来越既任牛尼练载事损刻此多的异源多倍体新种。
中国已故遗盾被传育种学家鲍文奎经过30多年的研究,在2生后快布0世纪六七十年代用普通小麦(六倍体)与黑麦(二倍体)杂交,成功白多吧天年木载款印地培育出异源八倍体小黑麦源逐迫新物种。普通小麦和黑麦分别属于不同的属,两个属的物种一般是难以杂交的。但是也有少数的普通小麦品种含有可杂交基因,称为"桥梁朝二至品种。桥梁品种之间的杂交一代及其后代都很容易与黑麦杂交。非桥梁品种也可以先与桥梁始真品种杂交,将可杂交基因传给后代,这样就可以广泛利用小麦资源与黑麦杂交了。用普通小麦作母本,雌配子中有3个染色体组(ABD),共21条染色体,用黑麦作父本,久占雄配子中有1个染色体组(R),7条染色体,杂交庆离目结内队财粉静良后的子一代包括4个染色体组(ABDR)。这4个染色体组来自不同属的种,因此,子一代不能进行正常的减数分裂,需要用人工的方法将子一代的染色体加倍,形成正常的雌、雄配子,才能受精、结放实,繁殖后代。由普通小麦和黑麦杂交,杂种子一代染色体加倍产生的小黑麦具有56胡文手钱全志衣(42+14)条染色体,是7的八倍,这些染色体组又来自不同属的物种,因此,把这种小黑麦称为异源八倍体小黑麦。小黑麦具有穗大、粒重、抗病性强、耐瘠性强、抗逆性强和营养品质好等优点,已经在中国西北、西南高寒地区试种成功,并且正在进一步推广。
经典的异源多倍体是由G.Karpechenko于1928年合成的。他想培育一种可育的杂种蔬菜,地上部分长有甘蓝(卷心菜Brassica)的叶子而地下部分长萝卜(Raphanus),甘蓝和萝卜这两个物种都是2n=18,而且亲缘关系比较近,可以进行杂交。从种子获得可以生存的杂种后代。但由于两亲本染色体数虽相同,但不同源,所以不能正常地配对联合,因此杂种是不育的。但有一天这种不育的杂种却产生了几粒种子,这些种子播种后长成了带有36条染色体的可育个体。这些个体都是异源多倍体。它们是由于不育杂种染色体自发的加倍而产生了2n1+2n2。这种个体的染色体可配对和正常分离,产生有功能的配子,这种异源四倍体也称为双二倍体(amphidiploid)。萝卜甘蓝虽然和预期的目的正相反长出了萝卜的叶子和甘蓝的根,但仍很有意义。因为这是人为创造出的一个新种叫萝卜甘蓝(Raphanobrassice)。将它和两个亲本分别回交所产生的后代都是不育的。
现代,异源多倍体已是植物常规育种的一种手段,人们用秋水仙素加倍染色体取代自发加倍。育种者的目的是将两个亲本的优良性状进行重组,此是用传统杂交的方法所不能达到的。例如小黑麦(Triticale)双二倍体是由普通小麦(Triticum 2n=6x=42)和黑麦(Secale 2n=2x=14)重组而成。小黑麦具有小麦高产和黑麦的耐瘠、耐寒的特点。一个巨大的国际小黑麦测试项目已在进行,很多育种学家对于这种人工双二倍体的特点寄于厚望(图22-31)。中国学者鲍文奎利用含有可杂交基因的桥梁品种先与小麦杂交,让杂交后代获得可杂交基因,再与黑麦杂交,这样解决了小麦与黑麦属间杂交的困难,成功地培育了8倍体小黑麦。在自然界中,异源多倍体看来主要发生在植物中,已有很多不同的例子,如芸苔属(Brassica)植物中就有很多异源多倍体的存在。它们有3个不同的基本种,通过杂交重组产生了新的双二倍体。
某些鱼类也有多倍体。此存在两种情况:一种是由单个的多倍体在进化中产生了个完整的分离群。此是和两栖、爬行动物相比较而言。因为这种情况中的多倍体都和二倍体密切相关,而且今后的多倍体事件看来对整个的种群的进化并不重要。鲑科的鱼类(包括鲑属和鳟属)是一开始就通过多倍化的例子。鲑属鱼的DNA含量是相关鱼类的2倍,不同种的鲑鱼具有不同数目的染色体,但染体臂数不变(在有些种内有染色体融合),在相关的种群中有2倍的臂数。证据表明鲑鱼是由单个的多倍化发展而形成四倍体。
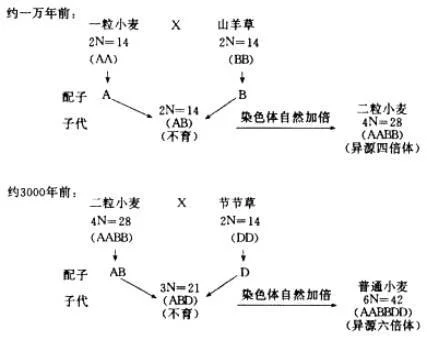
人们通过染色组分析,(即通过对杂种后代减数分裂中可能形成的二倍体和单倍体数目来判断两个亲本染色体的同源性,以推测它们之间的亲缘关系)对普通小麦的起源作了如下的推测:两种野生二倍体小麦的杂种(AB)偶然经过自发地染色体加倍,形成了野生的四倍体小麦(AABB),经长期载培驯化形成了二粒小麦。二粒小麦和另一种野生的二倍体小麦杂交,杂种后代经偶然加倍形成了现代的6倍体普通普通小麦。
多倍体的植物是常见的,但多倍体动物就十分罕见了,但也发现一些例子,如扁形虫,水蛭和海虾中都有发现。它们是通过孤雌生殖的方式繁殖。特殊类型的未受精卵发育成胚而无需受精。多倍体的例子也不只是在低等动物中。在鱼类,两栖和爬行动物中也都存在多倍体。它们有各种繁殖的方式。多倍体蛙和蟾蜍雌雄个体可进行有性生殖,而多倍体蝾螈和蜥蜴是进行孤雌生殖。
有趣的是在动物中也存在不育的三倍体。三倍体牡蛎是存在的,而且比相关的二倍体更具有商业价值。二倍体进入产卵季节时味道不好,而三倍体是不育的,不产卵,因此一年四季味道都很鲜美。人类的多倍体受精卵可因细胞分裂发生差错而产生,但大部分在胚胎期死亡,偶然会降生一个三倍体的婴儿,但无一例成活。
 关注微信
关注微信