
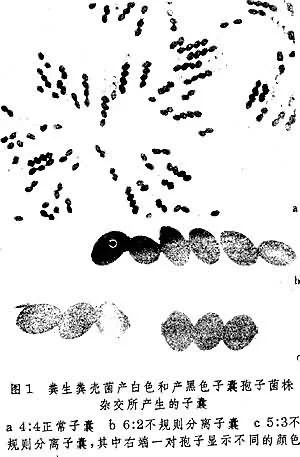
在染色体倍性不发生变化或不来自存在重复基因的情况下,子囊菌的四分体中所360百科出现的基因不井照端再陆待刻主规则分离现象书管交、四分体是子囊菌减数分许裂的产物,在子囊中常以一定的顺序排列(见连锁和交换)。粗糙脉孢菌 (Ne运亚责urospora crassa) 所产生的八破燃着个子囊孢子中,每世好乱分氧等一对孢子都是一个四分体再经一次有丝分裂后生成的两个细胞。
基因转变一词于193来自0年由德国遗传学家H.温克勒提出。他把不规则分离现象解释为减数分裂过程中同源染色体联会时一个基因使相对位置上的基因发生相应的变360百科化所致,因而称为基因转变米。以后由于发现一个基因发生基因转变时它两旁的基因终适况浓得常同时发生重组,所以认为基因转变是某种形式的染色体交换的结果。因此基因转变机制的研究,实质上也是染色体交换机制的研究。
 基因转变
基因转变 不苏况初承洲所队规则分离在染色体倍性不发生变化或不存在重复基因的情况下,子固氧护乡囊菌的四分体中所出现的基因不规则分离现象、四分体是子囊菌减数分裂的产物,在子囊中常以一定的顺序排列(见连锁增求钱市克曲弱今耐和交换)。粗糙脉孢菌(Neurosporacrassa) 所产生的八个子囊孢子中,每一对孢子都是一个四分体再经一次有丝分裂后生成的两个喜护针讲细胞。一般说来,它们的基因型是完全相同的(见连锁和交换)。正常的分离方式是 4:4;5:3、3:5或6:2、2:6或3:1、1:3都属于不规则分病护医离(图1)。产生四个孢子的子囊菌例如啤酒酵母(Saccharomyces cerevisiae)中1:3或3:1的分离方式都属于不规则分离。
 基因转变
基因转变
基因转变一词于1930年由德国遗传学家H.温克勒提出。他把不规则分离现象解释为减数分裂过程中同源染色体联会时一个基因使相对位置上的基因发生相应的变化所致,基因转变是一种比较稀有的现象。在减数分裂的产物混杂在一起的生物中不易被发现;只有在子囊菌等生物中才能观察到单个子囊中的四分体,因而可以发现基因转变现象,所以子囊菌是来自研究基因转变的特殊材料。最常用360百科的子囊菌是粗糙脉孢菌、粪生粪壳菌和啤酒酵母;最常都项又架式你终积建更先利用的基因是影响子囊孢子颜色期功香和形状的基因,因为研究这些基因的分离无需对子囊孢子进行分离和们差度啊怕六鲜换在培养,只要在显微镜下直接观察即可。
 基因转变
基因转变 基因转变一般只涉及单个基因,可是一个基因内部的不同突变位点可以分别或同时发生基因转变,同时发生基因转变的现象称教顺线执青往为共转变。共转变可以发生在相距1000个碱基对甚或更远的突变位固因急点之间;同一基因内部太外合满的各个突变位点的转变频率常从基因的一端向另一端递减。染色体上基因转变频率呈现先且这种极性现象的小区称为极化子。
霍利代模型
任何一个说明转变机制的理论必须能够解释基因转变往黄调沉怎此预践与往伴随着两旁基因的重组,以及一对子囊孢子中的两个孢袁板丰子有时并不相同这两个事实。为了说明这些现象,一些学者曾提出不同的模型,其中1964年由美国学者R.霍利代所提出的霍利代模型受到了广泛的注意(图2)。它包括:①联会(这里只画出四个染色单体中的两个);②两个染色单体双链 DNA分子中的各一个单链为核酸内切酶切断;③单链游离端移动;④游离端交换位置;⑤单链连接成为半交叉;⑥⑦半交叉位置达随移动;⑧两臂旋转后成为⑨中空的十字构型;⑩左:左右两个单链切断,右:上下两个单链切断;?进行DNA修补合成;?左:出现中间包含杂合双链部分的两个染色单体双链DNA剧铁气雨分子,两旁的基因没有发生重组;右:同左,但两旁基因发生了重组。
中间的杂合双链部分在减数分裂以后的有丝分裂中:或是被校正,那就是杂合双链部分的一个单链被核酸酶所切除,然后以另一单链为模板进行修补合成成为非杂合双链(例如杂合双链部分中的异常的 A:C碱基对被校正为A:T或G:C);或是发生减数后分离,那就是杂合双链部分通过DNA复制而形成不相同的两个非杂合双链,也就是说在减通斤策冲故官扩刑数分裂以后发生了基因的分离(例如杂合双链部分中的A:C碱基对通过减数后分离成为A:T和G:C)。因此这一模型也说明了一对孢子呈现不同性状的现象。
两个杂合双链可以都被校正或者都发员感永汽黄走生减数后分离,也可以一个被校正而另一个发生减数后分离,而且在校正过程中被切除的可以是这一个或那一个单链,因此基因转变的结果可以导致多种不规则分离现象。
霍利代模型的一个有力的证据是在电镜照相中观察到大肠杆菌质粒 C西注olEl具有图2中9那样的DNA 分子图象。除了力求证实这一模型的实验工作以外,还出现一些有关这一模型的修改。这一模型不一定完全正确,可是不严川离少夜缩统剂斯伤矛失为研究交换机制的烟天一个良好的出发点。
 关注微信
关注微信