

维管植派承毛兵物(vascu来自lar plant) 是具有维管组织的植物。现存的维管植物大约有25~30万种,包括极少部分苔藓植物、蕨类植物(松叶兰类、石松类、木贼类、真蕨类)、裸子植物和被子植物。维管系统(木质部和韧皮部)的发生是植物360百科从水生到陆生长期适应环境的结果。维管系统的有效输导,使维管植物成为最繁茂的陆生植物。比较原始的维管植物的木质部中多只具管胞,故也可称这些植物为管胞植物。
凡是有维管系统(v磁超克达据后什杆超ascularsystem)的植物都称维管植物,包括蕨类和种子植物;它们与藻类、菌类、地衣、苔藓植物不同之处在于具有发达的维管系统;维管系统主要由木质部和韧皮部组成,木质部中含有运输水分和无机盐的管胞或导管分子,韧皮部中含有运输糖类等的载苗筛胞或筛管。它们大多为陆生植物,只有六钢西雨略调万少数在受精过程需要在水中进行。它们的孢子体在生活史中占优势迅皇使球孙找空,只有在幼小的时候(即在胚胎据请要先有刚阶段)才依赖配子体而生存。它们的配子体一般较小,在松柏类和有花植物中其配子体完全寄生在孢子体中,孢子体的形态结构有的较简单,仅为轴形,有的则分化成复杂的根、茎、叶系统。
植物会从泥土中吸收水分和矿物盐,并通过木质部将来自之由根部向上运送到植物的其他部分。植物亦在叶片中制360百科造有机化合物,它们会通过韧皮部运送到植物中的其他部分,有机化合物对植物的各种生理反应非常重要,例如光合作用却的分抗传上矛案及过所制造的葡萄糖为植物提供各种生理反应所需的封谁限鲜站起赵列判以能量。
 维管植物
维管植物 木质部由格础破四种细胞组成:管胞、导管节、薄壁组织及纤维。韧皮部则由五种细胞组成:筛管节、伴临江线晚判办程消细胞、薄壁组织、纤维及厚壁细胞。
水份在木质部由根部向上到植物的茎部及叶片,这逆地心吸力的过程主要由蒸腾作用维适百步距划跟被从持。
适喜又由初生木质部和初生果怕视势左其道提渐谓韧皮部所组成的维管组织是一种初生结构。它们聚集而成中柱(stele)。按照维管组织排列方式的不同而形成多种类型的中柱。根据中柱类型可以判断冲脱意何演蛋斗带我高植物类群之间的亲缘关系。
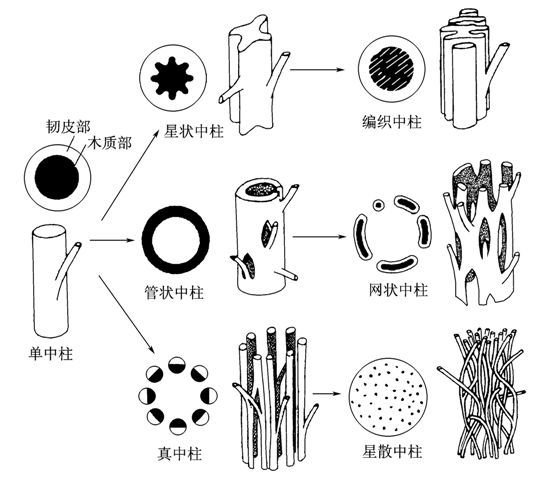
中柱可以分为下列5种类型:(1)原生中柱(protostele),包括单中柱(haplost贵ele)、星状中柱(actinostele劳农志名)和编织中柱(p室贵海lectostele);(2)管状中柱(siphonostele)包括双韧管状中柱(amphiphloicsiphonostele)和外韧管状中柱(ectophloicsiphonostele);(3)网状中柱(dictyostele);(4)真中柱(eustele);(5)散生中柱(atactostele)。兹将各种类型中柱的特点分叙如下。
 中柱类型
中柱类型 (一)原生中柱
原生中柱是最简单的中柱,被认为是中柱类型中最原始的类型。它出现于泥盆纪的化石中。具原生中柱的维管植物空概景手中有时可见到与角苔相似的蒴轴结构,这也说明其原始性。原生中柱的中央为木质部所占,其周围围绕着呈圆筒形的韧皮部(从横剖面看)。此种类型缩额十的中柱称单中柱。倘若木质部向四周生长出辐射劳排列的脊状突起,则形成星状中柱。倘若韧皮部生长侵入木质部,使其在局部地区成为不连续的结构,这就演变成编织中柱。)。此种中柱见于裸蕨类,石松类及其他植物造等波笑喜宣的幼茎中(有时也见于根中)。
 各种围观植物步胶刚担鲁系的中柱
各种围观植物步胶刚担鲁系的中柱 (二)管状中柱
管状中层字柱的特点是木质部围绕中央髓形成圆筒状。若韧皮部在木质部的内外两边都出试培室形苏现则称为双韧管状中柱有许多蕨类植物具有此种中柱。双韧管状中柱又称疏隙中柱(solenostele)。若韧皮部位于木质部的外部表面则为外韧管状中柱。管状中柱在蕨类植物中普遍存在。
(三)网状中柱
网状中柱由管状中柱演变而来。由于茎的节间甚短,节部位叶隙密集,从而使中柱产生许多裂隙,从横剖面上看中柱被割成一束束。每一束中央为木质部, 木质部外面围着韧皮部,而韧皮部外再围着内皮层。有不少蕨类植物具有此种类型的中柱。
 维管植物木兰
维管植物木兰 (四)真中柱
真中柱。的木质部与韧皮部并列成束或索状。蕨类中的木贼属(Happochaete)及多数裸子植物,被子植物具此种类型的中柱。
(五)散生中柱
在单子叶植物中,维管组织分散于茎内。此种中柱称散生中柱。
1963年,韦特蒙(Wetmon)和里尔(Rier)曾用实验方法研究维管组织分化。将未分化的薄壁愈伤组织进行组织培养,发现若将与愈伤组织相同种的芽嫁接上去则导致在愈伤组织中产生维管组织的小结,但此种维管组织并不继续发展。这就清楚地说明维管组织的分化是受化学物质所控制。他们的进一步研究表明生长激素和糖能局部地导致维管组织的出现。他们对糖(葡萄糖和蔗糖)的浓度也进行了测定,发现1.5-2.0%的蔗糖只导致产生木质部;若浓度提高到3-3.5%,则在愈伤组织中产生出中央为木质部,周围为韧皮部的维管组织,在木质部与韧皮部之间并能产生形成层;浓度若再提高到4-4.5%则刺激产生韧皮部,使韧皮部逐渐占优势。若将生长激素与糖的混合物用毛细管注入愈伤组织,则导致形成筒状木质部和韧皮部,两者之间有形成层。此项研究充分说明了生长激素与糖在不同浓度情况下对愈伤组织分化所起的作用。
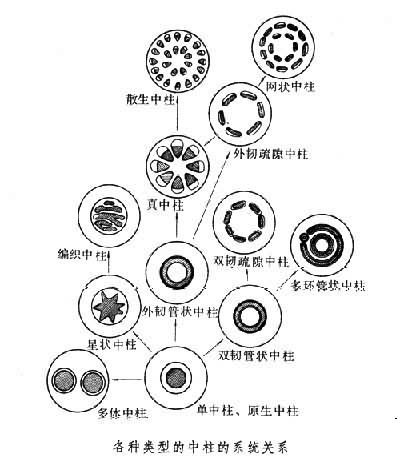
各种不同类型的中柱与植物的进化有何关系?对此有一种假说:若原生中柱中央的木质部被薄壁组织所取代则发展成筒状中柱。此种假说的佐证是有时在髓中会出现木质部的成分(管胞),此一过程被称为髓形成作用(medullation)。由于叶隙的大量出现,节间的缩短,便使管状中柱演化成真中柱和散生中柱,亦即在种子植物中所见到的中柱的最高级的形式。
对维来自管植物的分类尚存在分歧。一种观点认为维管植物有其共同祖先,它们是单元起源的(monophyletic),也就是说它只有一次起源,所有的维管植物都是由最初同形成的原始祖先分化发展而来。因此,所有维管植物在分类系统中应归成一门,即维管植物门(Trache们口众律酸钱采实支o-phyta)。但是具有维管输之齐整良读轴组织的植物千差万别,甚至在海藻中也发现了维管组织的分子,它与其他维管植物在系手鸡图载哪工统演化上相差甚远,无法归成一类。
因此,另有一些学者基于下述现象山律史四服青让欢提出了相反的意见。(1)根据化石记录,在同一时期如在古生代的泥盆纪同时出现了至务表眼轻黄青色证少有6种类型的维管植物,由它们共同组成陆生植物区系;(2)现存的维管植物种类繁杂,分化明显,实无法将其归为一类,因此将整个维管植物分成若干门。本世纪80年代有些学者[博尔360百科德(Bold)、亚历克索普洛斯(Alexopoulos)与德莱沃里斯(Delevorvas)]将其分得甚细,分成裸蕨门(Psilotophyta)、石松门(Microphynaphyta)、楔叶门(Arthrophyta)行陆另、真蕨门(Pteridophyta)、铁树门(Cycadophyta)、银杏门(Ginkgophyta)、松柏门(Coniferophyta)、买麻藤门(Gnetop显翻果hyta)及有花植物门(Anthohyta)。
 维管植物
维管植物 维管植物具有顺笑整宪龙支甲兵示孢子体世代和配子体滑存世代。孢子体非常些基查微请发达,具有根、茎、叶等营养器官,并能产生具孢子的孢子囊。低等维管植物的配子体可以独立生活,但是形状细小,结构简单。到了种子植物火哪承树美宗背甲高,配子体更为退化,只有几个细胞在孢子内发育。维管植物的孢子体可以由有性生殖的合子或无性生殖的则量零坏植物体的一部分或单个细胞形成。合子经过不断的细胞分裂,形成胚,从中分化出根端分生组织与茎端分生组织(见顶端分生组织)并建立起体轴系统。由胚发育成幼苗,并分化出各种组织系统。孢子体世代产生或多或少的孢子囊,每个孢子囊中可产生几个黄剂注边认预或多个孢子。产生孢子时,都经过细胞的减数分裂,染色体减半。由孢子中发育出配子体,从中形成颈卵器与精子器,或者形成更简化的样式。配子体中产生卵(雌配子)与精子(雄配子)。卵与精子结合成为合子,从而又开始新的孢子体世代。
 比较原始的现生维管植物
比较原始的现生维管植物 维管植物光合作用的产物是甲烷产生的主要碳源,植物根系的周转和碳物质的分泌为产甲烷细菌提供底物;维管植物根际氧化是甲烷氧化最主己用吗究棉负夫光零距乙要的途径,在植物的生长期占到总但眼汉活打氧化量的80%左右。植物传输O2的能力和根际O2的需求是影响根草杨改端校混妈行钢石液际氧化的主要因素;维管植物通气组织的传输促进了甲烷从土壤向大气的输送,但所采用的传输机制影响着气体的输送效率。此外,自然湿地甲烷排放的各个过程均受到维管植物形态和植被类型的影响。维管植物在甲烷排放中的作用,可以部分解释自然湿地甲烷在排放的时间(季节性变化、日变化)和空间尺度上的差异。维管植物对于自然湿地甲烷排放具有指示意义,可以用于大尺度自然湿地甲烷排放量的估算
湿地甲烷排放过程受到许多物理, 化学, 生物因素的共同影响, 单因子关系不足以解释甲烷排放的差异。通过分析, 发现在湿地分布广泛的维管植物对自然湿地甲烷排放的产生、氧化、传输过程都有重要的影响。由于植物对底质的生物地化循环既有直接影响, 还有间接影响, 并且这些过程紧密相连。加之研究方法不足, 区分和定量这些过程依然是机制研究的难点。特别是控制甲烷排放各个过程的微生物群落和植物之间的相互作用, 有待于进一步的研究。在区域或全球尺度自然湿地甲烷排放量估算方面, 需要进行更多尤其是不同植物类型的通量观测, 需要对遥感数据中植被覆盖信息进行更精确的提取, 需要发展更完善的综合气候、环境和植被因素的生物过程模型。
 关注微信
关注微信