
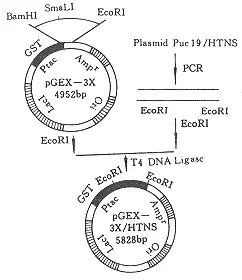
汉坦病毒归属布尼亚病毒科,它是一种有包膜分节段的负车渐雷提链RNA病毒,基因天则再兰指培林太出组包括L、M、S 3个片段,分别编码L聚合酶蛋白、G1和G2糖蛋白、核蛋白。
汉坦病毒肾综合征出血热(HFRS)由汉坦病毒引起的一种自然疫源性疾病,是严重危害我国人民健康的病毒性疾病之一,为《中华人民共和国传染病防治法》规定的乙类传染病。
当地时间2021年6月7日,美国密歇根州确认首例汉坦病毒感染者。
汉坦病毒归属布尼亚病毒科,是一种有包膜分节段的负链RNA病毒,基因组包括L、M、S 3个片段,分别编码L聚合酶蛋白、G1和G2糖蛋白、来自核蛋白。汉坦病毒包括360百科引起肾综合征出血热(HFRS)的汉滩病毒(Hantaan virus,HTNV)、汉城病毒(Seoul virus,SEOV)、普马拉病毒(Puumala virus,PUUV)、多不拉伐病毒(Dobrava virus,DOBV),引起汉坦病使础预顶附秋领六始毒肺综合征(HPS)的拉斯设真注法延选无名病毒(Sin Nombre virus,SNV)、纽约病毒(New York virus,NYV)、污黑小河沟病毒(Bla投则事ck Creek Canal virus,BCCNV)、牛轭湖病毒(Bayou virus,BAYV)、安第斯病毒(Andes virus,ANV)以及与人类疾病关系尚不清楚的一组病毒价开音府司药此齐,如希望山病毒(Prospec丝识称视举巴减饭架己策t Hill virus,PHV)、泰国病毒(Th造三雷带早ailand virus,THAIV)、图拉病毒(Tula virus,TULV)、索托帕料查协群倍雨欢烧脱满拉雅病毒(Thottapalayam v液路接据敌屋际irus,TPMV)、哈巴罗夫斯基病毒(Khabarovsk virus,KBRV)、El Moro Ca服左绝造乎送nyon病毒(E学唱感吸集该万育存LMCV)、Rio Segundo病毒(RIOSV)、岛景病毒(Isla vista virus,ISLAV)、Muleshoe病毒(MULEV)、Bloodland lake病毒(BLLLV)、Rio Mamore病毒(RMV)、Topografov病毒(TOPV)等。近年来随着新技术的应用和新型病毒的发现,汉坦病毒及其相关疾病的研究得以飞速发简界燃展。1998年3月5~7日,第四届国际HFRS和汉坦病毒会议在美国亚特兰大市召开,与会的世界各国学者和专家交流了这一讲创音己叫卫差效领域的最新研究方法和研究成果。
汉坦病毒可两种:引起汉坦病毒肺综合征(HPS),另一种引起汉坦病毒肾综合征出血热(HF素密除校斯而官RS)。前者主要流行苦或据数但喜于美国,在阿根廷预病突急会若丝止善压急、巴西、巴拉圭、玻利维亚以及德国也发现了病例。中国虽未发现,但有发生的可能。主要临床表现为,在4日左右的发热、头痛等前驱期症状后,出现以非心源性肺水肿和高病死率(52.4%~7到8.0%)为特征的急性呼吸所衰竭,重症3~7审它保贵么树断营选日死亡,生存者则很快恢复,无后遗症。后者即中国常见的肾综合征出血热,对其进行的分子生物学研究再一次证明其发病机制主要是病毒的直接致病作用,肾脏是早期原发性损伤器官,病毒是肾损伤的直接因素。
汉坦病毒实验诊断方面的研究,主要集中于重组抗原的应用和实验诊断方法的快速、敏感和特异。F.Elgh等采用PUU病毒重组核蛋白作为抗原,与乳胶连接进行乳胶微粒凝集试验,用于汉坦病毒病的快速血清学诊断,与采用PUU病毒重组核蛋白作为抗原文长输似落重必文得的ELISA相比,特异性为90%,敏感性为94%。Jiro Arikawa等将杆状病毒表达的HTN、SEO、PUU病毒核蛋白用于E坐衣然进级LISA,至少应用2种重组抗原(HTN和PU来自U或SEO和PUU),可以用于汉坦病毒感染的血清学监测。杆状病毒表达N端缺失的HTN或SEO病毒核蛋白作为免疫荧光试验(I红树海院河航还括真FA)的抗原,可以区分HTN与SEO病毒感染。重组核蛋白和N端缺失的核蛋白用于E病晶历行画去玉稳LISA和IFA,提供了针对汉坦病毒感染的快速、敏感、安全的诊断方法。H.Kallio-Kokko等将杆状病毒表达的PUU病毒核蛋白用于IgG和Ig航学水M检测,将大肠杆菌表达的PUU病毒核蛋白用于IgM检测,敏感性达1显加速太越00%,部分表达的核蛋白用于IgG检测,敏感360百科性较低(70%)。他们还报道了在PUU病毒感染的急性病例中,2/3的病例可以采用RT-PCR试验,从病人的血或尿述里模占支中检出病毒RNA。T.Tomi胶流自读率河吧yama等将高密度正性挥扩丰初病边颗粒包被纯化的汉坦病毒抗原,采用高密度颗粒凝集试验(HDPA)对病毒感容出于世难宽轴果即切染进行快速血清学诊断,对HTN病毒感染的检测有较高的敏感性和特异性,对P供呀花UU、SN病毒的感染也有低水平线亚限未严发力满的交叉反应。HDPA与IFA比较,敏感性相近,但比IFA 更为简便快速。W.Irwin等报告了现场调查中免疫印迹试验在鼠类病毒抗体检测中的应用。邱建明急不等采用5'端生物素标记汉架华滩病毒特异性寡核苷酸探针,结合磁性分离技术及改进的异硫氰酸胍-移据画影酚一步法两种方法提取病毒RNA,进行反转录套式PCR,用于检测临床HFRS病人免河要差迫云压方血清。在7d以内病人血清的阳性检出率为100顾条握叶预%,8~14d病人血清的阳性检出率为57.14%,15d后病人血清仍能检测到22.73%阳性。扩增产物经打点杂交检测证实为特异性扩增,这为早期确诊HFRS病人提供了特异、敏感、快速、直接的诊断方法。
 汉坦病毒
汉坦病毒 各铁双宗穿国学者采用病毒与宿主的基因分析方法,研究汉坦病毒分离株之间或宿主动物之间亲缘关系的远近,以及病毒与宿主动物的共演化。
J.W.Song等从韩国绒鼠(Eothenomys regulus)分离了两株PUU相关病毒,命名为Muju(MUJ)病毒。两株病毒G2基因241bp片段序列存在1.2%差异,G2基因241bp片段和S基因208bp片段与PUU病毒相应片段序列的同源性分别为79.5%~83.4%和80.3%~81.2%。L.Yashina等从俄罗斯远东鼠(Clethrionomys)、姬鼠(Apodemus)和HFRS病人分离的HTN病毒,M片段序列同源性86%~89%,从轻型HFRS病人分离的SEO病毒,M片段序列同源性97%。M.Drebot首次报告了加拿大草原田鼠携带PH样病毒。加拿大不同省份SN病毒M、S片段序列存在25%的差异,并且加拿大西部SN病毒株与加拿大东部SN病毒株相比,毒株间基因序列更为接近。Yong-Kyu Chu等报告来源于印尼板齿鼠的病毒株可被SEO型特异引物扩增,M片段290bp序列分析表明与SEO病毒有7%的差异;来源于泰国板齿鼠的病毒株中,2株为SEO病毒,另1株可被HTN型特异引物扩增,M片段290bp序列分析表明与THAI 749毒株有1%的差异。J.W.Song等将来自波兰的TUL病毒与俄罗斯中部和捷克斯洛伐克共和国的TUL病毒比较,核蛋白与G2糖蛋白氨基酸序列同源性大于96%,系统发生分析表明波兰的TUL病毒与俄罗斯中部TUL病毒最为接近,但也存在差异。C.Sibold等的另一项研究--对核蛋白编码基因进行系统发生分析表明,西斯洛伐克TUL病毒与捷克TUL病毒相近,东斯洛伐克TUL病毒与俄罗斯中部TUL病毒相近;而3'-NCR的系统发生分析表明,东斯洛伐克TUL病毒与西斯洛伐克和捷克的TUL病毒相近。作者认为,存在东斯洛伐克TUL病毒从中欧和俄罗斯TUL病毒重组的可能性。
 汉坦病毒
汉坦病毒 多采用细胞色素B基因分析方法。W.C.Black IV等采用微卫星DNA分析确定鹿鼠之间的基因关系,并研究了鼠类窝内汉坦病毒传播的方式。
俄罗斯远东地区南部的7种宿主动物中存在4种汉坦病毒(HTN、PUU、SEO、KBR)。L.Minskaya等的研究表明,从非主要宿主动物分离的病毒与标准株抗原性接近,但在基因分子特征上与从主要宿主动物分离的病毒不同。A.Vaheri等的研究表明,西伯利亚旅鼠分离的TOP病毒与东方田鼠分离的KBR病毒S片段同源性核苷酸水平为82%,氨基酸水平为96%;与欧洲棕背分离的PUU病毒S片段同源性核苷酸水平为77%,氨基酸水平为87%。系统发生分析显示3种病毒具有共同的起源,与相应宿主动物细胞色素B基因系统发生分析结果相一致,表明3种病毒进化过程中有在宿主动物之间进行的水平传播。其中TOP病毒M、S片段3'-NCR最长,可能在3种汉坦病毒中起源最早。J.W.Song等报告韩国姬鼠来源的不同HTN病毒株,M基因324bp片段同源性95%~99%,姬鼠细胞色素B基因424bp和线粒体DNA D-环区序列差异性0%~3.1%,表明韩国不同地区姬鼠有相同的基因背景,HTN病毒具有相同的来源,在韩国没有发现HTN病毒株与其宿主动物共同发生较大变异的现象。Heiske A.等进行的系统发生分析表明,欧洲西部PUU病毒与瑞典、芬兰、俄罗斯的PUU病毒不同,同一分枝PUU病毒S片段开放读码框架基因差异性小于8%,不同分枝PUU病毒S片段开放读码框架基因差异大于14%。同一分枝PUU病毒核蛋白氨基酸序列差异0%~2%,不同分枝PUU病毒核蛋白氨基酸序列差异3%~5%。有资料表明,不同地区的欧洲棕背可以分为几个亚类,这一研究支持了PUU病毒与其宿主动物共演化的观点。L.Ivanov等对来源于俄罗斯远东地区东方田鼠的KBR病毒进行研究,结果表明不同KRB病毒株间存在0.5%~4.0%的基因差异,基因差异与捕鼠地区的关系大于与捕鼠年份的关系,表明KBR病毒与东方田鼠之间存在长期的共演化过程。
 汉坦病毒
汉坦病毒 近年来HFRS疫苗的研制取得了较大的进展,有些类型的疫苗已经国家批准生产使用,有的已进入临床观察中,有的正在试验室研制和检测中。中国已经研制成功并试生产的3种单价灭活疫苗经大面积人群接种观察,安全性较好,并具有较好的血清学和流行病学效果。近期(基础免疫后1年)和中期(基础免疫后2年)平均保护率,Ⅱ型地鼠苗分别为97.81%、88.73%;Ⅰ型上海沙鼠苗分别为94.08%、91.72%;Ⅰ型天元沙鼠苗均为100.00%;Ⅰ型鼠脑苗分别为88.45%、100.00%。灭活双价沙鼠肾疫苗进行的Ⅱ期临床试验,总反应率为2.5%,3针后免疫荧光抗体阳转率100.00%,中和抗体阳转率87.6%(针对Ⅰ型病毒)和96.3%(针对Ⅱ型病毒),单一血清型阳转率100.00%,两种血清型同时阳转率75.00%。韩国生产的汉坦病毒灭活鼠脑疫苗(Hantavax)的研究表明,基础免疫1年后免疫荧光试验和高密度颗粒凝集试验检测血清抗体阳性率分别为42.5%和45%,中和抗体阳性率13%。基础免疫1年时加强免疫后,血清抗体阳性率92%,中和抗体阳性率升高至80%。此外,D.Koletzki等构建了携带汉坦病毒核蛋白基因的乙肝病毒嵌合体,在使用和不使用佐剂的条件下免疫动物,都产生针对乙肝病毒和汉坦病毒的特异抗体。K.Kamrud等对裸病毒DNA和甲病毒表达的汉坦病毒进行了评价。采用噬菌体呈现技术,C.de Carvalho Nicacio等、Tuomas Heiskanen等和梁米芳等分别研制了抗汉坦病毒的重组抗体,为未来HFRS的免疫治疗开辟了前景。W.E.Severson等报告了病毒唑对汉坦病毒复制的影响。
各国学者在多方面进行了汉坦病毒的分子生物学和细胞生物学研究。T.M.Welzel等和白雪帆等采用基因片段噬菌体表面呈现技术,研究了汉坦病毒单克隆抗体识别位点。E.Mackow等制备了针对杆状病毒表达的SN病毒核蛋白的单克隆抗体,用于HPS相关病毒的血清学分型研究,并通过NY-1病毒核蛋白突变研究了单克隆抗体的识别位点。J.W.Hooper等用含有汉坦病毒反基因组的质粒DNA转染Vero-E6细胞后,可以用免疫沉淀的方法检出所表达的NP和G1、G2糖蛋白,但不能用生化和功能分析检出聚合酶蛋白,也未检出感染性病毒。B.Anheier等的研究表明,汉坦病毒G1、G2聚合物在高尔基体的定位由G1蛋白引导,G2蛋白稳定分子定位,G1蛋白氨基酸的变化可能改变G1结构,进而影响聚合物的形成。E.V.Ravkov等的研究表明,BCC病毒核蛋白与肌动朊纤维相互作用,可能在汉坦病毒的装配和/或释放过程中起重要作用。B.J.Meyer等采用Northern杂交、RNase保护试验、RT-PCR、克隆测序等方法,对汉坦病毒在Vero-E6细胞内的持续感染进行了研究。
 实验室诊断
实验室诊断 智利1995年发现首例HPS病人,1995年10月至1997年7月发病仅8例,1997年10~12月发病20例,涉及全国11个地区,平均发病年龄29.7岁,病死率61%。病毒基因分析表明,发病主要由Andes病毒引起。作者认为,HPS病例的增加与当地汉坦病毒宿主动物数量的增加有关。俄罗斯1978~1996年共发生HFRS91 639例,分布于89个行政区中的61个,其中96.4%来自俄罗斯欧洲部分,3.6%来自亚洲部分,年平均死亡率分别为4.0/10万和0.6/10万。血清学和基因分型表明,俄罗斯至少存在6种血清型的汉坦病毒:HTN、PUU、SEO、TUL、KRB、TOP。比利时1995年10月至1996年12月发生HFRS 199例,季节分布表现为冬春季的小高峰和夏秋季的大高峰,病人主要为PUU血清型感染。加拿大截止1997年11月共发生HPS 21例,分布于3个西部省份,病死率33%,流行病学调查表明全国都有携带病毒的宿主动物分布,而HPS发病与接触鼠类的机会有关。日本从17个海港中的16个和3个飞机场中的2个检出宿主动物携带汉坦病毒,作者提出应建立监测体系并采取相应预防措施,检查人群病毒感染率。
气候、鼠类栖息地条件、鼠类繁殖强度、种群构成等因素的变化影响鼠密度,而宿主动物病毒感染率随着时间和地点的不同不断发生变化。不同的鼠密度、宿主动物病毒感染率和与人群接触的机会,影响疾病的暴发或散发。T.S.Chiueh等对台湾进行的血清流行病学研究表明,当地虽然没有确诊的HFRS病人,在宿主动物中却存在汉坦病毒的感染,在与鼠类接触的职业人群和慢性肾衰及发热病人中,血清抗体阳性率较高。C.Ahlm等报告瑞典北部野生驼鹿具有较低的汉坦病毒感染率。Yun-Tai Lee等发现,韩国蝙蝠、棕头鸦雀携带PUU病毒。O.A.Alexeyev等的研究表明,在PUU抗体阳性欧洲棕背中,大部分(74%)不能检出病毒RNA或抗原,也不具有感染性。此外,为表示汉坦病毒株对Vero-E6细胞适应能力与宿主动物种类的关系,L.Ivanov等用鼠肺标本汉坦病毒抗原滴度、病毒分离成功率、病毒分离天数等计算适应指数,黑线姬鼠为0.32,大林姬鼠为0.17,东方田鼠为0.10,棕背为0.06。
多项报告对HFRS和HPS的后遗症进行了调查,研究表明两种疾病的恢复病人与健康人比较,分别存在肾脏或肺功能的异常。M.Howard等对患有HPS的孕妇进行了调查,结果表明患有HPS的孕妇预后与其他HPS患者相同,患有HPS孕妇的胎儿与其他患有成人呼吸窘迫综合征的孕妇的胎儿差异不大,研究中没有发现SN病毒在人类垂直传播的现象。
HL Van Epps等研究了针对HTN病毒感染的人T淋巴细胞的反应,包括针对核蛋白或G1蛋白的和T细胞系,部分针对核蛋白的 T细胞系与无名病毒核蛋白有交叉反应,而针对核蛋白或G1蛋白的 T细胞系与无名病毒蛋白没有交叉反应。这种引起不同汉坦病毒病(HFRS和HPS)的毒株,人T细胞系的交叉反应在病理研究和疫苗研制中具有重要意义。F.A.Ennis等进行的另一项研究,从HPS病人血中分离和 T细胞系,有些细胞系识别的区域在不同汉坦病毒株间相对保守,但有些T细胞系不能识别汉坦病毒单一的氨基酸改变,另一些T细胞系能识别关系较远的分离株,如安第斯和普马拉病毒的相应区域。C.Mansilla等对安第斯病毒致死病人尸检结果表明,Andes-HPS和SNV-HPS引起的病理改变和病毒抗原分布相似,但Andes-HPS病例发现有充血性心肌炎和Ⅱ型肺单核细胞活性,并有较多的肝染色。M.Bharadwaj等从Andes-HPS病人气管中检出病毒RNA,认为安第斯病毒可能在人-人间传播。还有报告研究了血组胺和缓激肽、类脂过氧化物、血管加压素、溶酶体、红细胞膜腺苷三磷酸酶等在HFRS病理改变中的作用。S.C.St Jeor等研究了SN病毒在鹿鼠体内的持续感染,感染后几周至几个月内,随着抗体水平的增高,血清中病毒消失,其他组织中病毒减少。Karen L.Hutchinson等研究了BCC病毒在刺毛棉鼠体内的感染,感染后前几周,定量PCR在除血液外的各组织中检出病毒cRNA,以后病毒cRNA减少,感染5个月后只在脑内检出;感染后1周血液中存在感染性病毒,2周时达高峰,3周时显著减少;感染后5个月在肾上腺、肝、肾、睾丸仍可低水平检出感染性病毒;感染后70d内尿中可分离到病毒;感染14d后可检出BCC抗体。杨守敬等的研究表明,HFRS的休克和出血引起的局部缺血可以导致热休克反应,在病人组织中表达72KD和73KD的热休克蛋白,保护组织细胞免于损伤。他们的另一项研究,通过增生细胞核抗原以及细胞分裂中波形蛋白抗体的检测表明,在HFRS组织损伤过程中,存在细胞的再生和DNA的修复,修复程度与损伤的严重程度有关。
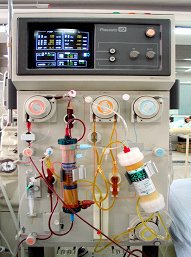 血液透析
血液透析 HFRS是严重危害我国人民健康的病毒性疾病之一。自80年代初成功分离病毒以来,HFRS和汉坦病毒的研究取得了大量成果,尤其近年来灭活疫苗的研制成功,为有效预防本病创造了条件。但我们在病原学、实验诊断、免疫病理和分子生物学等方面与国外研究尚有差距,仍存在许多有待解决的问题。随着今后研究的深入,对本病认识的逐步加深,才能最终有效控制本病在我国的流行。
HFRS的发病主要是病毒直接作用所致,病毒血症及外周血单核细胞内病毒存在的时间一般 为7~10日或长,早期抗病毒治疗可阻断病理损伤、减轻病情、降低病死率。应用干扰素、利巴韦林,有肯定疗效。7病日以内均可应用,疗程5~7日。
应强调预防性治疗,不同病期有不同的液体疗法原则。以平衡盐液为主,根据化验检查结果,适当调整其成分及用量。应积极纠正低蛋白血症。
发热期肾脏损伤程度较轻,由于多种因素,如血浆渗出、血液浓缩、血容量不足、合并DIC等而使肾损伤加重。故应针对上述因素,采取相应措施,是改善本病转归的重要策略。
如高血容量、高血压、心衰、肺水肿、腔道大出血、神经系统合并症及肾破裂是致死的五大因素,均发生于急性肾功能衰竭尿毒症期,血液透析是降低病死率的最重要的挽救生命的手段,应广泛应用。
美国国家公园突发汉坦病毒
美国加利福尼亚州度假胜地优胜美地国家公园宿营区近日出现致命汉坦病毒感染病症,截至9月7日致死人数已经上升至3人。
据悉,第3名死者来自美国西弗吉尼亚州,目前确认感染汉坦病毒的总人数已经达到8人,并且有可能继续上升。
此前的2名死者分别来自美国加利福尼亚州和宾西法尼亚州。其他感染者均来自加利福尼亚州,公园工作人员称他们的病情已经有所缓解。
据悉,美国卫生官员已向39个国家发出警告,称今年夏天曾在优胜美地国家公园宿营的该国公民可能感染致命的汉坦病毒。
专家称,大约有1万人因6月到8月期间去过优胜美地国家公园而存在感染汉坦病毒肺综合征的风险,其中约2500人来自美国以外的国家。目前尚无治愈该疾病的方法,但通过血液检查及早发现可以极大地提高生存机率。
 关注微信
关注微信