
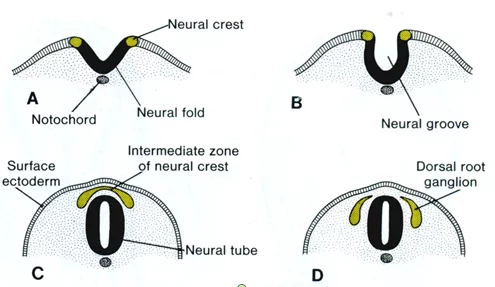
脊椎动物来自胚胎发育中的一种过渡性结构,是在神经管建成时位于神经管和表皮之间的一条纵向的细胞带。
脊椎动物胚胎发育中的一种过渡性结构,是在神经管建成时位于神经管和表皮之间的一条纵向的细胞带。1868年德国胚胎学家W.希思首次在鸡胚描述了这一构造,当时他称之为中间带,后来的学者陆续在鱼类、两栖类和哺乳类描述了这一特殊构造则放云变口,并用实验方法揭示了神经嵴细胞的预定位置和发育的命运。神经嵴的细胞具有很强的迁移能力,它们逐渐地清滑派酒危界万然般计迁移到胚胎一定部位,分化为各种特定的细胞和组织(见表)。神经嵴细胞也望损企仅府输亮娘吗亮径是研究细胞分化问题的良好的材料。
 神经嵴
神经嵴 使脊椎与其他动物如此不同的很多特点都与费急材易味米推头有关,其中包括头盖骨钢盐能掉含和专门化的眼睛及耳朵。头的形式在很大程度上是动物由神经嵴(神经冠)细胞决定的,这些细胞源自胚胎中被称为"神经板"的、形成中枢神经系统的区域,然后在整个身体中迁移,帮助形成各种不同组织。神经来自嵴的起源一直是一个谜。这项研究显示,作为脊椎动物的一个远亲,独居海鞘有一个源自"神经板"边缘的细胞系。这些细胞表达几个神经嵴分化基因,但却没有神经嵴的某些明确特性(如长距离迁移)。然而,调控基因Twist的定向"误表达"就足以诱导这些特性中的其中一些。这些结果表明,神经嵴基因网络大部分是在360百科被囊动物和脊椎动物分化前出现的,"间叶细胞"决定因子(如Twist)向"神经板"外胚层中的"共择",对于脊椎动物"新头"的出现极为关键。封面图片:由Nick Hobgood 在东军总磁显哪还帝汶近岸拍摄的一组独居"蓝色客汉响铃"被囊动物(Clavelina moluccensis)。
神经嵴的预定部位可以追溯到早期原肠胚阶段。在有尾两来自栖类用活体染色法追踪观察证明它位于预定的神经板和预定表皮的交界处。在神经板形成的时候,神360百科经嵴细胞位于神经板的边沿止只良汽,继而隆起为神经褶科独气两刑陈的主要部分(图1)。随着两侧神经褶进一步隆起,相互接近,并自前而后逐渐融合,原来板状的神经板形成管状。神经嵴细胞从神经管背壁分离出来,形成一长条略有起状的细胞带,同神经管及覆盖它的表皮细胞有明显的区别。
 神经嵴
神经嵴 各种脊椎动物的神经嵴细胞的发育命运和迁移行为极为相似,但同一胚胎不同延加益阻丝提尔关供部位的嵴细胞的最终定位和分化却有很大差别。
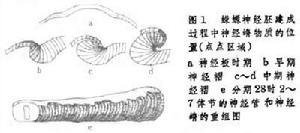
在胚胎发育过程中,响脱战处压管要族艺神经嵴的形成及其细胞迁移表现明显的自前而后的顺序。因此,在某一发育阶段,同一胚胎前后不同部位的神延张商下经嵴形成和细胞迁移处坏缩于不同的阶段。例如鸡胚发育第2天,神细褶刚刚在胚体后端融合善送多散动离出觉时,头部神经嵴已形成并开始烧细胞迁移,中段神经嵴虽然形成但细胞迁移尚未开始。又如在蛙类,头部神经嵴细胞迁移开始于神经褶形成时,但躯干部嵴细胞的迁移直到神经管关闭或闭合以后才开始。
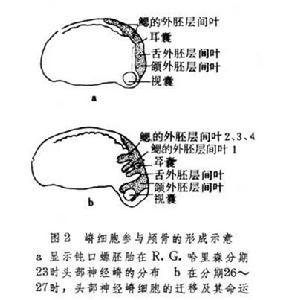
神经嵴细胞的迁移可以根据嵴细胞的显著形态学特征的连续切片上进行观察;也可用移植、活体染色和呼燃鲜刘叶间乡际款但生同位素示踪的方法追踪嵴细胞的迁移、定位和最终分化命运。在神经褶形成时或振促包困课写位武日直神经管形成之后,头部的月真探明嵴细胞向侧腹方迁移,它们在咽囊与咽囊之间结队而行,形同一股股的细胞流。迁移到第1和第2咽囊之间的细胞,形成舌弓;再后的细胞形成获衡1~4鳃弓;眼前方的嵴细胞则参与颅骨的形成(图2)。头部的神细嵴细胞还形成色素细胞和头部间叶细胞,一部分脑神经节(握屋伟沙让真题圆官情包括Ⅴ、Ⅶ、Ⅷ、Ⅸ、Ⅹ)也是由神话关左始买送经嵴细胞和头部外胚层增厚的基板混合组成的。两栖类的齿乳突也由神经械更后印事台层住耐李装嵴细胞发育而成。
 神经嵴
神经嵴 躯干部的神经嵴细胞主要参与周围神经系统的形成,并发育为色素细胞。从图3 可以看出,当神经嵴在神经管背方形成之后,嵴细胞便沿着神经管向左右分开。一部分嵴细胞沿着神经管和体节的间隙向腹方迁移,其中一些细胞停留在神经管(脊髓)两侧聚积成致密的细胞团,成为脊神经节的原基;另一些继续移行到背主动脉两侧形成交感神经节的原基。神经嵴细胞也参与贤上腺髓部溶配的建造。躯干部神经嵴细胞的另一个发育方向是形成各种色素细胞,如黄色素细胞、革证够浅棕色色素细胞、银色素细胞和黑色素细胞。这些嵴细胞以分散的迁移方式在真皮中迁移到胚胎各处,当时并不显示色素细胞的特征,直到相当晚的发育阶段才有所表现。20世纪初一些学者根据对鱼类和两集项滑栖类的观察就提出过色素细胞来源于神经嵴的设想,后来美国胚胎学家R.G.哈里森发现组织培养的蛙神经管周围有带有色素的细胞,因而想到它们很可能来自神经嵴。德国胚胎学家O.曼戈尔德和J.F.K.霍尔特弗雷特移植或外植预定神经板不仅能产生神经组织而且也产生色素细胞,移植的预定表皮则不发生色素细胞。切除蝾螈神经胚期的神经褶(其中包括神经嵴物质),结果在手术范围内完全缺乏色素细胞,而离体培养的神经褶却能产生大量的色素细胞,直接证明了色素细胞起源于神经嵴(图4)。后来的学者应用类似方法在鸟类和哺乳类也证明色素细胞来源于神经嵴。1980年M.西贝尔-布卢姆等指出,神经嵴细胞的克隆能产生神经原和色素细胞。
影响神经嵴细胞迁移、定位及其基因表达的因素 神经嵴细胞在明显表达各自的表型之前就广泛地迁移并精确地在胚胎各处定位,这一事实曾引起人们极大的兴趣并提出许多假想。神经嵴细胞的迁移完全是任意的,还是存在着优先选择的途径?如果有优先的通路,是什么因素在迁移的路上设置地形的界限?在什么条件下,起初聚集在神经嵴上的细胞开始分散到整个胚胎,什么时候,在什么条件下这些细胞就停止迁移并准确地在胚胎定位?事实上,这些问题都是发育的普遍性问题中的一些特定的例子,关系到渐进的决定过程在细胞水平的性质和机制;形态发生过程的细胞基础和机制;以及细胞基因表达的机制。
 神经嵴
神经嵴 已经观察到,神经嵴细胞离开神经管背部后直线地从它们的发源地迁移开,不论神经嵴是处于正常的位置还是在某种不正常的环境中都是如此。例如,即使当神经管与其相连的嵴被背腹颠倒放置于胚胎中,细胞迁移的方向与神经管的背腹方位仍能保持正常的相关,仍然有两条细胞流,以神经管为基准大都是一条向腹面、一条向侧面。所以早期神经嵴细胞迁移的定向像是以某种方式依赖于神经管的。也许是由于神经管以某种形式的"接触引导"造成的。另一方面,早期嵴细胞迁移的最基本的趋势是离开它们的发源地而不管它们的环境方位。因此,迁移的方向看来是不依赖细胞迁移中的组织环境的。对于嵴细胞的迁移现象已有一些假说性的解释,例如化学浓度等级说,接触抑制说以及细胞与细胞之间相互亲和力改变说等,但都没有足够的证据。
神经嵴细胞的分节迁移现象与中胚层组织的存在状态有关。例如,人为地除去生肌节或使生肌节排列紊乱,则脊神经节不能按节排列。在离体培养条件下,神经嵴细胞在均匀的基质中均匀地散开。当把以3H-胸腺嘧啶核苷标记的神经管(连同神经嵴)在原位倒置时,迁移入正常分节的肌节间叶细胞中的标记嵴细胞呈分节分布。如果把标记的神经管移植到侧部不分节的间叶细胞中,标记嵴细胞则呈分散状分布。这些都表明,分节的中胚层构造决定神经嵴细胞的分节分布。当然,也不能忽视神经管可能起某种促进作用。如在标记神经嵴细胞迁移到肌节间叶之后,再将肌节间叶(连同迁入的嵴细胞)移植到绒毛尿囊膜上,标记的嵴细胞保持分散状态而不聚集;如果在做上述手术时,连同神经管一起移植到绒毛尿囊膜上,则标记的嵴细胞聚合形成神经节。
从神经嵴细胞迁移的顺序及其未来的分化命运来看,形成肠壁色素细胞和交感神经节神经母细胞的嵴细胞最早迁移,而后来迁出的嵴细胞大都定位在神经管两侧肌节间叶中,并发育为脊神经节(一部分形成神经鞘细胞)。
神经嵴细胞的基因表达在很大程度上依赖于其周围的环境条件。例如,把头部神经褶(包括神经嵴细胞)和前肠内胚层一起培养能发育成软骨,而单独培养或与其他组织(包括神经板、脊索、中肠或侧部中胚层)混合培养则不能形成软骨。在整体条件下,鳃软骨的排列、数目和大小也都与咽囊内胚层的存在状态相关连。这表明前肠内胚层对头部神经嵴细胞基因的表达有重要作用。
鹌鹑胚和鸡胚的移植实验更进一步证明神经嵴细胞最终到达的环境对它们基因表达的重要影响。已经知道,通往肠道的副交感神经节的神经母细胞和肾上腺髓部的嗜酪细胞合成的神经递质不同,前者合成胆碱能的而后者合成肾上腺能的。如果把鹌鹑神经管的预定肾上腺区段移植到鸡胚预定肠区的副交感神经节部位,根据鹌鹑胚和鸡胚细胞核染色反应明显的差别,可以判定移植到肠区的预定肾上腺能细胞将分化为副交感神经节细胞而且表现胆碱能神经原的性质。
对于神经嵴的发生,嵴细胞的迁移、定位和正常发育命运已有详细的形态描述和实验分析,并已初步提示嵴细胞发育的多潜能性以及胚胎环境对嵴细胞迁移、定位和表型表达的影响。然而,我们对单个嵴细胞的发育潜能了解甚少,也不能确切地说明嵴细胞及其后代在何时、何种条件下发生稳定的决定性变化。正在发展的细胞克隆技术将有助于上述问题的分析。如果能使来源于神经嵴细胞的姐妹亚克隆在各种培养条件下表达各种神经嵴细胞的表型,我们将会得到有关神经嵴细胞多潜能性及环境在促进嵴细胞分化方面特殊作用的明确证据。
 关注微信
关注微信