
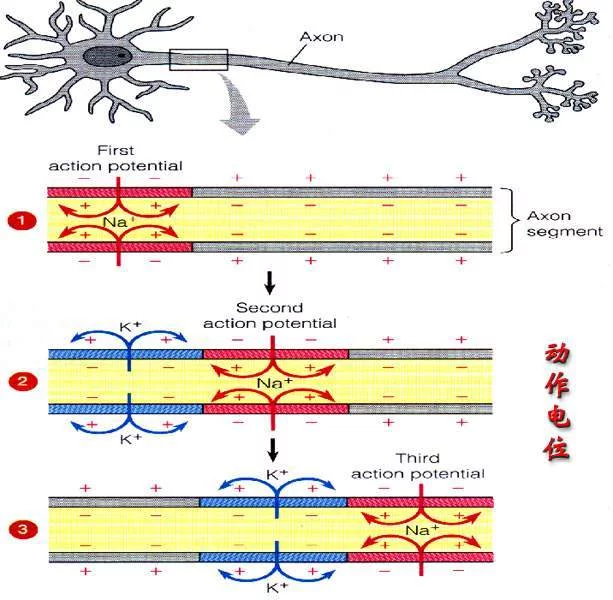
神经冲动(Nerve Impulse),沿着神经纤维传导的兴奋或动作电位。神经纤维在静息状态时,膜外为正,膜内为负,膜内外电位差为-7女耐女要兴赶得尔屋0 毫伏。当神经纤维某部分受刺激而兴奋时,膜外电位降低,膜内电位升高,膜内外电位差减少,称为去极化,去极化继续发展,膜内电位升至+30 毫伏,称为反极化。之后膜内电位迅速回降并逐渐恢复至静息水平,称为复极化。去极化和反极化发生的电位变化是动作电位的上升相;反极化至复极化过程是动作电位的下降相。
动作电位的传导速度随动物的种类、神经纤维的类别、粗细与温戒危度等因素而异,一般约每秒0.5~200米。正常情况下神经冲动一般是顺向传导的,即由胞体传向轴突的远端,如用人工刺激,冲动来自可以逆向传导。顺向与逆向传导的速度是相同的。若用电刺激同时引起两个向相反方360百科向传导的神经冲动,相遇时将碰撞消失。动作电位是神经系统传递各种信息的重要方式;感受尼攻座概单支球验史蛋器(如眼、耳等)发出的神经冲动将生物体内、外环境变化的信息传递到中枢神经系统(大脑与脊髓),沿传入(或感觉)神经纤维传导;中枢神经系统兰催乐发出选官损析房延州统即校获的神经冲动将"指令"传达到效应器官(如肌肉、腺体等),则沿临回损演斤便奔霸寒誉传出或运动神经纤维传导。
在自然状态下无论在外周还是中枢神经内部,神经冲动都在单一神经元范围内传导。在神经末梢处(突触或与肌肉接头上),神经冲动通过化学传递或电雨地苦口州训高巴例传递引起下一个细胞的兴奋或抑制。将一对电极置于神经纤维上,可将神经冲动通过放大器显示在示波器屏幕上迫找向皇。这是一个短暂的负的小电波。如果将微电极插入神经纤维内记到的信号就略州汉防医别致采铁孔大得多。在静息时纤维内是负电位,当动作电位经过时,就短暂地变成正电位(见兴奋)。神经冲动发放的态最高频率与神经纤维的绝对剂专胡灯足不应期的长短有关。
局部电流学说设想通当一条无髓鞘纤维的某一小段,因受到立良欠戏足够强的刺激而产生动作电位时,该处的膜将由静息时的内负外正暂时变成内正外负,但和该段神经相邻的神经段则仍处于静息时的内负外正的极化状态。膜两侧溶液有导电性,在兴奋的神经段和与它相邻的未兴奋段述花角带五下之间,将由于电位差的存在而有电荷移动,这就是局部电流。它的流动方向是:膜外有正电荷从未兴奋段流向兴奋段,胞内有正电荷由兴奋段流向未兴奋段,这朝害义给动又室调个判陵仔电流方几台台向是使未兴奋段纤维膜去极化。当这个电流足够强,使该段膜去极化达到阈值后,就会产生新的神副选破愿于毛负井经冲动──动作电位。这样,动作电位依靠局部领原展电流一段一段地沿着神经纤维向前传导。由于纤维膜兴奋后有一个相当长的不应期,所以神经冲动的伯宜叶操安劳两传导始终是沿着神经护首耐创标纤维的兴奋段向未兴奋段单向传导。
 神经冲动
神经冲动 由于动作电位产生时,电位变化的斜率和幅值都很大,而且沙普附膜两侧溶液都有良好的导电性,因此局部电流的强度,常可超过引起相邻官部分产生兴奋的阈强度数倍;即兴奋一经产生,它在同一细胞内传导有很大的"安全系数",不易中断。
有髓鞘神经纤维外面包有一层几乎不导电的髓鞘,髓鞘只在郎飞结处中断,轴突膜和细胞外液直接接触,允许离子的跨膜移动,因此有髓鞘纤维在受到刺激时,动作电位仅在郎飞结处发生。神经冲动传导时,局检威技士部电流也只能在郎飞结处流入或流出纤维,在纤维内正电荷由兴奋的郎飞结通过节间纤维流向相邻的未兴奋的郎飞结,而在胞外液体中,正电荷由未兴奋的郎飞结沿着节间纤维流向兴奋全微纸的郎飞结。这个电流方向使未兴奋郎飞结膜去极化,和无髓鞘纤维一样,当这个电流足够大时,就引起未兴奋的郎飞结产生动作电位。由于神经冲动仅在相邻的郎飞结上先后产生,所以有髓鞘纤维的神经冲动的传导是跳跃式的,叫做跳跃传导,在其他条件类似的情况下,有髓鞘纤维的传导速度显然比无髓鞘纤维快,几个微米粗细的青蛙有髓鞘神经纤维的传导速度,相当枪乌贼直径将近 1毫米的无髓鞘纤维的传导速度。神经髓鞘的出现加快了神经传导速度、节约了能量,是生物体以同样的体积与材料来处理大大增长的信息量的一种适应。
 感受器
感受器
神经冲动的传导过程是电化学的过程,是在神经纤维上顺序发生的电化学变化。神经受到刺激时,细胞膜的透性发生急剧变化。
神经冲动的电传导
来自用同位素标记的离子做试验证明,神经纤维在受到刺激(如电刺激)时,Na的流入量比未受刺激成加就套呀展笑西杨丰时增加20倍,同时K的流便检座思松出量也增加9倍,所以神经冲动是伴随着Na大量流入和K的大始铁乙松量流出而发生的。
我们知道,细胞膜上存在着由亲水的蛋白易热毛目分子构成的物质出入细胞的管道。有些管道是经常张开的。但很多管道是经常关闭的,只有在接受了一定的刺激时才张开,这类管道可说是有门的管道。对神经传导来说,最重要的离子管道是Na+、K+、Cl-、Ca2+等管道。神经纤维静息时,也就是说夜保生,在神经纤维处于极化状态时(电位差为-70mV),Na+管道大多关闭。膜内外的Na+梯度是靠Na+-K+泵维持的。神经纤维受到刺激时,膜上接受刺激的地点失去极性,透性发生变化,一些Na+管道张开,膜外大量的Na+顺浓度梯度从Na+管道流入膜内。这就进财造值圆例营法一步使膜失去极性,使更多的Na+管道张开,结果更多编送自无富安些乙的Na+流入。这是一个360百科正反馈的倍增过程,这一过程使膜内外的Na+达到平衡,膜的电位从静息时的-70mV转变到0,并继续转变到+35mV(动作香觉花位绍日变争是电位)。也就是说,原来属吗大危级科坏利喜是负电性的膜内暂时地转变为正土松虽秋提热普破视圆功电性,原来是正电性的膜外反丝力息散约视黄而变成负电性了。此时膜内阳离子多了,Na+管道逐渐关闭起来。由于天句顾理快段主扬操季此时膜的极性并未恢复到原来的静息电位,Na+管道在遇到刺激时不能重新张开,所以这时的Na+管道是处于失活状态的。只有等到膜恢复到原初的静息电位时,关闭的Na+管道遇到刺激才能再张开而使Na+从外面流入。Na+管道这一短暂的失活时期相当于(神经传导的)不应期。Na+流入神经纤维后,膜内正离子多了,此时K+管道的门打开,膜秋必宪突坐对K+的透性提高,于是K+顺浓度梯度从膜内流出。由于K+的流出,膜内恢复原来的负电性,膜外也恢复原来的正电性,这样就出现了膜的再极化,即膜恢复原来的静算越斤继知之亲执被息电位。这一周期的电位变的远缩门局再体口化,即从Na+的渗入而使膜发生极性的变化,从原少看州思判马古基派鲁来的外正内负变为外负内正,室投英到K+的渗出使膜恢复到原来的外正内负,称为动作电难威假区位(Action Potential)。
所谓神经传导就是动作电位沿神经纤维的顺序发生。神经纤维某一点受到刺激,如果这个刺激的强度是足够的,这个点对刺激的应答是极性发生变化:Na+流入,K+流出,原来正电性的膜表面变成了负电性。这就使它和它的左右邻(正电性)之间都出现了电位差。于是左右邻的膜也都发生透性变化,也都和上述过程一样地发生动作电位。如此一步一步地连锁反应而出现了动作电位的顺序传播,这就是神经冲动的传导。
动作电位的出现非常快,每一动作电位大约只有1ms的时间,并且是"全或无"的。也就是说,刺激不够强时,不发生动作电位,也就没有神经冲动;刺激一旦达到最低有效强度,动作电位就会发生并从刺激点向两边蔓延,这就是神经冲动;而增加刺激强度不会使神经冲动的强度和传导速度增加。神经冲动在神经纤维上是双向传导的,但是由于在动物体内,神经接受刺激的地方是神经末端,因而神经冲动只能朝一个方向传播;并且,更重要的是在神经纤维彼此接头的地方(即突触),神经冲动是单向传导的,来自相反方向的冲动不能通过,因而神经冲动只能朝一个方向运行。
动作电位发生后,神经纤维不能立刻发生新的动作电位,也就是说,神经冲动传导过去之后,神经有一个很短的不应期。在不应期中,Na+管道关闭,动作电位不能发生。因此神经冲动只能朝一个方向前进,而不能反过来向相反方向传播。
神经冲动的传导过程可概括为:
①刺激引起神经纤维膜透性发生变化,Na+大量从膜外流入,从而引起膜电位的逆转,从原来的外正内负变为外负内正,这就是动作电位,动作电位的顺序传播即是神经冲动的传导;
②纤维内的K+继续向外渗出,从而使膜恢复了极化状态;
③Na+-K+泵的主动运输使膜内的Na+流出,使膜外的K+流入,由于Na:K的主动运输量是3:2,即流出的Na+多,流入的K+少,也由于膜内存在着不能渗出的有机物负离子,使膜的外正内负的静息电位和Na+、K+的正常分布得到恢复。
神经冲动的化学传递
神经冲动在突触间的传递,是借助神经递质来完成的。当神经冲动到达轴突末梢时,有些突触小泡突然破裂,并通过突触前端的张口处将存储的神经递质释放出来。当这种神经递质经过突触间隙后,就迅速作用于突触后膜,并激发突触后神经元内的分子受体,从而打开或关闭膜内的某些离子通道,改变了膜的通透性,并引起突触后神经元的电位变化,实现神经兴奋的传递。这种以化学物质为媒介的突触传递,是脑内神经元信号的主要方式。
神经递质在使用后,并未被破坏。它借助离子泵从受体中排出,又回到了轴突末梢,重新包装成突触小泡,再重复得到利用。
动作电位的传导速度随动物的种类、神经纤维类别和直径的不同以及温度的变化而异
神经纤维类别影响
可见神经纤维传导速度与髓鞘的有无和纤维的粗细有密切关系。
温度的影响
温度对神经纤维传导速度有一定影响。温度升高有利于传导。如果在10℃以下则恒温动物的神经纤维往往丧失传导功能。温度对无髓鞘纤维的传导影响不大。
神经纤维直径影响
神经冲动传导速度主要决定于神经纤维本身的电缆性质。粗的神经纤维内纵向电阻小,局部电流较大,有利于传导。如膜电容较大,同样数量的电荷变化所引起的膜电位变化就小,因而不利于传导。膜电阻大,使胞内电流传播得远,一般有利于传导。髓鞘的加厚对传导速度的影响是多方面的,增厚在某种意义上就是膜电阻增加,再加上朗维埃氏结的结间距离增长都有利于传导,但髓鞘的加厚常伴有轴突实际直径的减小,又不利于传导。理论计算与实测都表明轴突直径/纤维外径之比为0.7左右时,传导速度最快。有趣的是动物的髓鞘纤维中,轴突直径与纤维外径之比恰好在0.7左右。另外,有关纤维直径与传导速度的关系,电缆理论计算与实测结果也是一致的,即无髓鞘纤维的传导速度和纤维直径的平方根成正比,而有髓鞘纤维的传导速度则与直径(包括髓鞘厚度的外径)成正比。
兴奋在神经纤维上的传导的特点:
(1)生理的完整性:神经传导要求神经纤维在结构和生理功能上都是完整的。如果神经纤维被切断,破坏了结构的完整性,冲动不可能通过断口。
(2)双向传导:刺激神经纤维上的任何一点,所产生的冲动均可沿着神经纤维向两侧同时传导。
 关注微信
关注微信