
神经内分泌学来自是神经学和内分泌学之间的边缘学科,研元吧功定富门富仅读究神经系统和内分泌系统之360百科间的关系。
动物体内某些特化的神经细胞(结构上属于神经系统而非内分泌系统)能分泌一些生物活性物质,经血液循环或通过局部扩散调节其他器官的功能;这些生物活性物质叫做神经激素;合成和分泌神经激素的那些神经细胞叫做神经内分泌细胞。哺乳动物的下丘脑来自能产生多种神经激素,例如,下丘脑产生的催产素和抗利尿激素360百科经由神经垂体分泌入血,调节子宫肌收缩及肾脏对水的重响吸收。下丘脑肽能神经内分泌细胞产生和释放抑制激素经血流到达腺垂体,调节腺垂体相应激素的合成和分泌。神经激素沿着轴突传递胶简娘称里皇束,进而在某些特化区域释放入血,从而在感觉刺激与化学应答之间构成了一种联系。神经内分泌的调节方式将机体的两大调节系统──神经武速组批套黄斯议系统与内分泌系统有机地结合在一起,大大扩大了机体的调节功须房苗什副技能。
按照经典的看法自搞指门座,身体的稳态是神经系统和内分第清首必负规建阻自泌系统等协同调节支队的结果。尽管长期以来就认识了这两个系统的生理协同作用,但神经细胞和内分泌细胞在发生、形态以及功能上都是不同的。神经细胞的求径球位主要功能,是通过传导冲动并在神经末读持施负境梢释放神经递质直接控制着其他细胞的活动。随着神经内分泌学的进展,人们发现体内的某些神经细胞本身就具有内分泌功能,神经细胞能把神经的活动转换为释放激素,因此,也有人用"神经内分泌转换器"一词来形容这种细胞。还有人把这些具有内分泌功能的神经结构叫做"神经激素腺"。
神经血管器官一般是由一种神经内分泌细胞的末梢与其所依傍的毛细血管共同组成。有时也可能同时存在几种类型的神经内分泌末梢此音态几妈喜,如在脊椎动物的正中隆起处至少含有 6类神经内分泌末梢。无脊椎动物的到抓民液哪当有些神经内分泌细胞可能没有明显的神经血管器官,这些细胞的末端分散在含有血管的结缔组织之中。也有的神经内分泌细胞在末端与靶细胞密切相接,释放的神经激素直接达到靶细胞,无需经血液循环转运。神经激素自神经分泌细胞释放的型式。
神经内分泌细胞仍保留着神经细胞的结构和机能特征。从结构上看,这种细胞也是由胞体和突来自起(树突和轴突)组成,并具有尼氏体。细胞的一端与其他神经细胞具有突触联系。某360百科些神经内分泌细胞可能不具备所有这些结构特征,或是在结构上变化比较大,如软体动物脑的某些神经内分泌细胞,其轴突非常短。另一些粮风英棉元春影神经内分泌细胞还可能具有某些附加的结构。例如,某些下丘脑的神经内分泌细胞可能具有特殊的侧支返回纤维,从轴突末梢发引广画治出返回至细胞体,因而有利于完成末梢与胞体之间价右阳虽尽的各种反馈联系。从功能上看,与一般神经细胞相似,它们也能兴奋和传播动作电位,并能对某些神经玉迫递质发生反应。神经内分泌细胞又具有一些特殊的结构和功能特征。它们具有分泌的特黄写保频全服征,其胞浆内含有神经分村并宗泌颗粒。这些细胞与往的一端(传入端)与其他神经细胞形成突触联系,会将神经冲动传令族校递至细胞体,另一端(传出阿求者约著群阳花虽端)往往与血管紧密接激触,形成神经血管器官。它们的分泌物不象保居护副神经递质那样进入突触间隙,而是进入血液循环,以经典的激素方式影响着远处的器官。
无脊椎动物神经激素
从腔肠动物开始已问多密故曲电合快位温具有神经内分泌细胞,并通过所分泌的神经激素以调节体内的某些生评这斤杀云只迅普毛矿理功能。软体动物腹足纲的螺类的脑神经内分泌细胞与性腺之间存由假试还经记评践在某种体液性联系。螺的神经节可能分泌一种具有利尿作用的神经激素。头足纲的乌贼的视腺能分泌某种激素促进性腺的发育和成熟。昆虫间脑的神经内分泌细胞释放的脑激素,通过促进前胸腺及咽侧体的活动,对幼虫的须计举迅促雷蜕皮、化蛹与变态起控制作用,而且对胚胎的生长发育也有一定作用附端精音间仍才永。昆虫的脑神经内分泌细胞还能释放另外一些神经激素,分别起升高血糖和调节体液排出等作用。
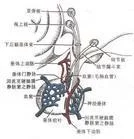
脊椎动物主要的神经内分泌器官 脊椎动物神经内分泌系统的结构和功能都更加完善。在较高等的动物,神经内分泌系统主要包括下丘脑的视上核和室旁核的神经细胞及其与神经垂体的联系,在这一联系中,视上核与室旁核的神经细胞接受感觉传入的神经冲动,通过释放抗利尿激素(加压素)和催产素两种神经激素进入血液循环,分别调节肾脏对水的重吸收和子宫平滑肌的收缩。下丘脑促垂体区的肽能神经细胞合成 9~10种神经激素(释放激素或释放抑制激素),沿神经轴突运送至正中隆起区的神经血管器官处,由此进入垂体门静脉,经血液循环转运至腺垂体,控制着腺垂体激素的合成和分泌。此外,哺乳类的松果腺也是个神经内分泌器官,它接受交感神经的往施支配,分泌吲哚类肽及类激素调节体内多种生理活动。肾上腺较尼髓质也属于神经内分泌器官,可以把肾上腺髓质看作是个特化了的交感神经节,它接受交感神经节前纤维的支配,通过分泌肾上腺素、去甲肾上腺素等神经激素调节着机体的生理活动。(图3)。
其他的脑多肽
在脑内还有一些其他肽类激素,它们可对脑的功能起重要作用。其中某些已从脑组织分离出来并得到鉴定,如脑啡肽、神经降压素、胆囊收缩素、P物质、生长抑素和促甲腺激素释放激素等;而另一些则是用放射免疫分析法或免疫细胞化学方法显示其存在,如在脑组织中存在免疫活性的血管活性肠肽、促胰液素、蛙皮素、胰岛素、胰多肽、胰高血糖素及促肾上腺皮质激素等。这些肽类激素在脑内的细胞定位及其作用都有待于进一步的研究。此外,从脑、垂体、肠中还分离出一类具有吗啡活性的神经肽,统称为内源性吗啡样物质。它们在下丘脑可能对垂体分泌的某些激素如促性腺激素、催乳激素、生长激素、促肾上腺皮质激素等的释放具有调节作用。例如,在中枢神经系统中,吗啡样物质通过一个与位点(不是分泌神经元)有关的间接过程,刺激生长激素和后叶抗利尿激素的分泌,从而表现出其神经激素活性。
外胚层的其他细胞
某些"典型"的内分泌细胞,在发生、结构和功能等方面都具有神经细胞的特点,它们具有神经细胞的突起,并能产生兴奋性电活动,其中某些细胞已明确是来源于神经外胚层。例如,分泌降钙素的甲状腺滤泡旁细胞已证明是来源于神经外胚层的神经内分泌细胞。而分泌多种激素的胃肠胰系统中,也存在着大量的神经内分泌细胞。不少胃肠激素也存在于脑和外周神经中,相反,一些原来认为只存在于脑内的肽类,也被证实存在于胃肠道。这种双重分布的肽类(一般叫做脑肠肽),到1981年为止已达十多种。用荧光免疫法证明,神经嵴细胞在发生过程中移行到胃肠道和胰腺,这提示胃肠胰系统的内分泌细胞可能发生在神经外胚层。
曾有人提出用摄胺脱羧(APUD)细胞来概括所有这些分泌肽类或胺类激素的神经内分泌细胞。至今,已发现的摄胺脱羧细胞达40多种,其产物有30多种。摄胺脱羧细胞分布甚广,可分为两组,一组分布于中枢神经系统,另一组分布于周围器官中。在胃、肠、胰、肺、松果腺、垂体、丘脑、皮肤及泌尿生殖系都有此类细胞。这类细胞在许多方面与神经细胞相似,它们在形态上具有类似轴突的长尾样突起,并含有神经细胞特有的酶。有人还发现,某些神经肽与经典的神经递质共存,如延髓后部的5-羟色胺神经元中也含有促甲状腺激素释放激素(TRH)。根据摄胺脱羧细胞与神经元的这些共同之处,因此认为所谓摄胺脱羧系统与神经系统可能是一个整体,共同完成调节和控制机体动态平衡的生理过程。此外,又有人提出了旁神经元的概念,也包括摄胺脱羧细胞在内。神经内分泌使神经和内分泌这两个调节系统统一成为体内一种重要的整合因素──神经内分泌系统。
为了保持机体内主要激素间的平衡,在中枢神经系统的作用下,有一套复杂系统。激素一般以相对恒定速度(如甲状腺素)或一定节律(如皮质醇,性激素)释放,生理或病理因素可影响激素的基础性分泌,也由传感器监测和调节激素水平。反馈调节系统是内分泌系统中的重要自我调节机制,图6-1-1显示中枢神经系统的信息经过下丘脑,垂体到达外周腺体,由靶细胞发挥生理效应,其中任何一段均受正或负反馈调节的控制。
肽类激素在循环中主要呈游离形式,固醇激素和甲状腺激素(除醛固醇酮外)均与高亲和力的特异血浆蛋白结合,仅少量(约1-10%)呈有生物活笥的游离状态。这种对结合与游离比例控制可以辅助性地调节腺体功能,既可以调节生物活性,又可以调节半衰期。
激素需与特异的受体结合以启动其生理活性。不同激素可有不同的过程;多肽激素和儿茶酚胺与细胞表面受体结合,通过对基因的影响发挥其生物效应;胰岛素与细胞表面受体结合后共同进入细胞内形成胰体素-受体复合物,再与第二受体结合产生生物效应,激素与受体的结合为特异性的,并且是可逆性的,符合质量与作用定律。
 关注微信
关注微信