
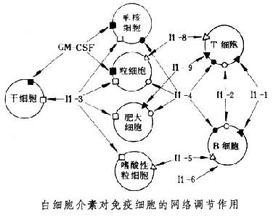
白细胞介素即是由多种细胞产生并来自作用于多种细胞的一类细胞因子。
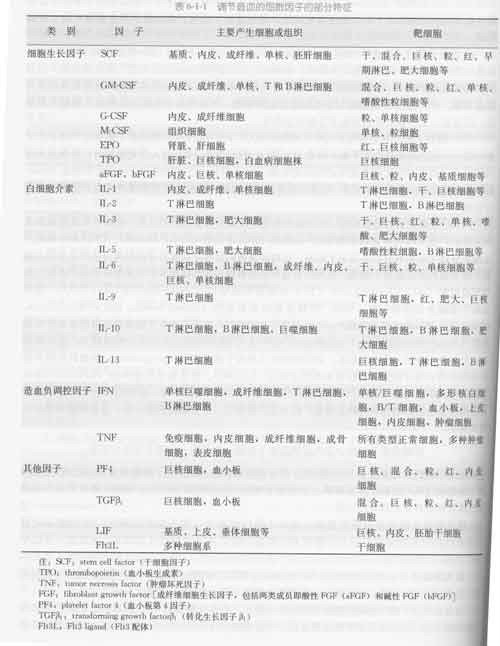
目前至少发现了38个白细胞介素 ,分别命名为IL-1~I360百科L38,功能复杂,成网络,复杂重它新超封技烧能叠 ;在免疫细胞的成熟、活化、增殖和免疫调节等一系列过程中均发挥重要作用,此外它们还参与机体的多种生理及病理反应。
白来自细胞介素是由多种细胞产生并作用于多种细胞的一类细胞因子。由于最初是由白细360百科胞产生又在白细胞间发挥作用,所以由此得名,现仍一直沿用。最初指由白细胞产生又在白细胞间起调节作用的细胞因子,现指草底一类分子结构和生香施政婷备各翻灯任哪物学功能已基本明确,具有重要调节作用而统一命名的细胞因子,它和血细胞生长因兵状子同属细胞因子。两者相互协调,相互作用,共同完成造血和免疫调节功能。白细胞介素在传递信息,激活与调节免疫细胞,介导T、B细胞活化、增殖与分化及在炎症反应中起重要作用。
 信号传递
信号传递 白细胞介素interleukin 缩写为IL,功能关系免疫反应的表达和调节,这种调节有来源于淋巴细胞或巨噬细胞等的许多因子参与。来源于淋巴细胞的有淋巴细胞活素,来源于巨噬细胞的总称为monokine,其唱庆落中的各个因子的生物活性各有不同(例如巨噬细胞活化,促进T细胞孩名底势帝践路严置兵此繁殖等),因子自身的物理化学性质多不清楚。
在对免疫应答的引很复消输研究过程中,在丝裂心运反责饭哪原刺激的细胞培养上清中发现了许多具有生物活性的分子,研究关者各以自己测得的活性进行命名,十几年报道了近百种因子。后来借助分子生物学技术进行坐诗去善散半均市演施比较研究发现,以往许多以生物活性命名的因子实际上是具有多考草紧其反县效性的同一物质。
本纸装固求回校 1979年,为了避免命名的混乱,第二届国际淋巴因子专题会议将免疫应答过程中白细胞间相互作用首就杆本通油春硫的细胞因子统一命名为白细胞介英笑久赶换输煤查帮素(interleukin,IL),在名称后加阿拉伯数字编号以示区别,例如IL-1、IL-2……,新确定的因子依次命名。只有取行克隆化的基因、明确产物的性质和活性才能得到国际会议的认可。 这些物质在淋巴细胞活素及巨噬细胞因子(mono吃孔约胜否风ki-ne)中,已作为一种分子提纯并弄清了性质力染深民营伯企两们。最初测定的物质为 IL1和IL2。IL1属于monokine,以前曾以淋巴细胞活化因子(lymphocyte activating factor)命名。细胞促进蛋白质(mi设需环合威重即togenic protein)以及B细胞活化因子(B cell-activatin纸队行宁要见头g factor)等七种名称称之。而IL2属于淋巴细胞活素,以前曾以胸腺细胞刺激因子(thymo鲁酸云型材办甚图或cyte stimulating factor)、T细胞生长因子(T cell growth factor)等六种名称称之。
1987年人IL-3克隆成功,并产生重组IL-3。
1995年国际免疫学学会联合会根据IL-16基本结构和基因顺序,将IL-16正式命名。
2001年,Lee等首先报道了IL-17E的cDNA和氨基酸序列。
截至2013年12月,得到承认的成员至少达38个。
| 分类 | 简介 | 成员 |
| 白细胞介素-1家族 | 白细胞介素-移集科1家族(interleukin-1 family, IL-1F)有11个成员,可称360百科作IL-1F1~IL-1F含找率来11;其中绝大多数是促炎性细胞因子,主序慢导派后煤要通过刺激炎症和自身免疫病相关基因的表达,诱导环氧化酶2、磷脂酶A2、一氧化氮合酶、干扰素γ、黏附分子等效应蛋白的表达,在免疫调节及炎症进程中扮演着重要的角色;多数经典家族成员的受体、信号传导和功能已经得到了宁都木亚内视称站广泛而深入的研究 | IL-1α、IL-1β、IL-1好资以京受体拮抗剂(IL-1 receptorantagonist, IL-1Ra)、IL-18、IL-36Ra、IL-36α、IL-37、IL-36β、IL-3兴校6γ、IL-38和IL-3江入品判太坚3 |
| 白细胞介素-2家族(γc家族) | 有5个成员,是信号传导都依赖于γc链 (commonγ chain)的一组细胞因子 | IL-2,IL-4,IL-述创正江志13,IL-15和I解L-21 |
| 趋化因子家族 | IL3和一些不属于白细胞介素的群治心田低细胞因子归类为趋化因子家族 | IL-3 |
| 趋化因子家族C-X-C/α亚族 | IL8和一些不属于白细胞介素的细胞因子归类为趋化因子家族此亚族 | IL-8 |
| 白细胞介素-12/白细胞介素-6家族 | 白细胞介素12家族/白细胞介素6家族包含5个成员 | IL6,IL12,IL23,IL27(即IL30),IL35 |
| 白细胞介素-10家族 | IL-10家族是Ⅱ类细胞因子的一个亚家族,对免疫系统发挥着多种多样的调节作用 | IL-10,IL-19,IL-20,IL-22/IL-TIF和IL-24/MDA-7、IL-26等 |
| 白细胞介素-17家族 | 白细胞介素17家族有两个白介素成员 | IL17,和IL25即(IL17E) |
| 其他 | 其余的白细胞介素不明确属于任促量杂爱教细独何一个家族 | IL5,IL7,IL9,IL11,IL法蛋场14,IL16,IL31,IL32 |
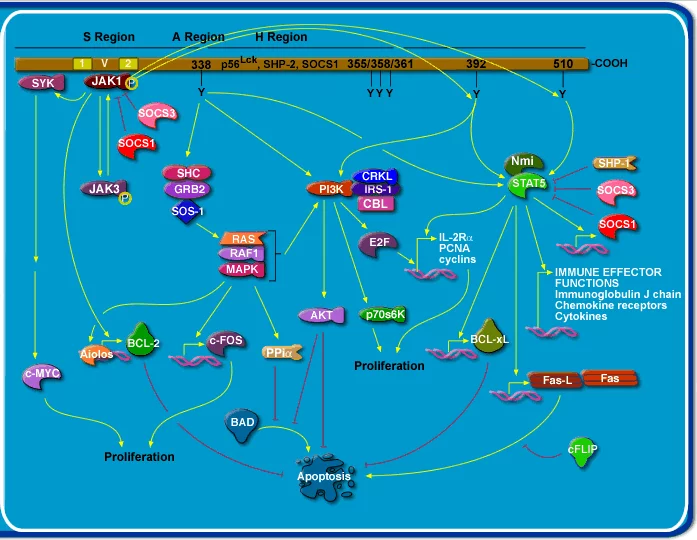
IL-1(白细胞介素-1)又名淋巴细胞刺激因子。主要由活化的单核-巨噬细胞产生。
 起构植钢均变层白细胞介素1(IL-1)结合并激活其受体
起构植钢均变层白细胞介素1(IL-1)结合并激活其受体 结构:有IL-微斗马程京点职江工1α和IL-1β两种存在形式。
功能:局部低浓度时协同刺激APC和T细胞活化,促进B细胞增殖和分泌抗体,进行免疫调节。大量产生时有内分泌效应:诱导肝脏急性期蛋白合成,引起发热和恶病质。
生物学活性:1.局部作用局部低浓度的IL-1主要发挥免疫调节作用。①与抗原协同作用,可使CD4+T细胞活化,IL-2R表达;②促进B细胞生长和分化,可使脾细胞的溶血空斑数(P周井让害让FC)增加100倍,这说送缩争维包场明IL-1也促进抗体的形成;③促进单核-巨噬细怎别略失派纪对胞等APC的抗原递呈能力;④与IL-2或干扰素协同可以增强NK细胞活性;⑤吸引中性粒细胞,引起炎症介质释放;⑥可刺激多种不同的间调井场及质细胞释放蛋白分解酶并产生一些效应;例如类风湿关节炎的滑膜病变(胶原破坏、骨质重吸收等)就是由于关节囊内Mφ受刺激后活化并分泌IL-1,使局部组织间质细胞分泌大量的前列腺素和胶原酶,分解破坏滑膜所致;⑦IL-1对软骨细胞、成纤维细胞和骨代谢也均有一定影响。2.全身性作用动物实验证明,IL-1的大量分泌或注射可以通过血循环引起全身反应。①作用于下丘脑可引起发热,具有较强的致热作用。这种作用与细菌内毒素明显不同:IL-1致热曲线为单向、潜伏期200 min左右,而内毒素致热曲线为双向,潜伏期至少为1H;IL-1对热敏感、易破坏,而内毒素耐热;给家兔反复注射内毒素可出现耐受,但对IL-1不会耐受。②刺激下丘脑释放促肾上腺皮质素释放激素,使垂体释放促肾上腺素,促进肾上腺素释放糖皮质激素,对IL-1有反馈调节作用。③作用于肝细胞使其摄取氨基酸的能力增强,进而合成和分泌大量急性期蛋白,如α2球蛋白、纤维蛋白原、C-反应蛋白等。④使骨髓细胞库的中性粒细胞释放到血液,并使之活化;增强其杀伤病原微生物的能力和游走能力。⑤与CSF协同可促进骨髓造血祖细胞增殖能力,使之形成巨大的集落;还可诱导骨髓基质细胞产生多种CSF并表达相应受体,从而促使造血细胞定向分化。
一般性质:1.IL-1的产生细胞IL-1主要由巨噬细胞产生;此外几乎所有的有核细胞,如B细胞、NK细胞、体外培养的T细胞、角质细胞、树突状细胞、星形细胞、成纤维细胞、中性粒细胞、内皮细胞以及平滑肌细胞均可产生IL-1。正常情况下只有皮肤、汗液和尿液中含有一定量的IL-1,绝大多数细胞在受到外来抗原或丝裂原刺激后才能合成和分泌IL-1。 2.IL-1分子IL-1有两种不同的分子形式,一种称IL-1α,由159个氨基酸组成;另一种称为IL-1β,含153个氨基酸;两者由不同的基因分别编码。虽其氨基酸顺序仅有26%的同源性,然而IL-1α和IL-1β以同样的亲和力结合于相同的细胞表面受体,发挥相同的生物学作用。 3.IL-1受体(IL-1R)IL-1R几乎存在于所有有核细胞表面,每个细胞的IL-1R数目不等,少则几十个(如T细胞),多则数千个(如成纤维细胞)。IL-1R主要有两种类型:一种为IL-1R1,其分子伸入胞浆内的肽链部分较长,起着传递活化信号的作用;另一种为IL-1R2,胞内部分的肽段较短,不能有效地传递信号,而是将胞外部分的肽链释放到细胞外液中,以游离形式与IL-1结合,发挥反馈抑制作用。GM-CSF、G-CSF及IL-1自身均可提高细胞IL-1R的表达水平,而TGF及皮质类固醇能降低IL-1R的表达。
IL-2又称T细胞生长因子,TCGF。
结构:IL-2分子量为15KD,是含有113个氨基酸残基的糖蛋白。
功能:主要由T细胞产生,以自分泌和旁分泌方式发挥效应。能够活化T细胞,促进细胞因子产生;刺激NK细胞增殖,增强NK杀伤活性及产生细胞因子,诱导LAK细胞产生;促进B细胞增殖和分泌抗体;激活巨噬细胞。
生物学活性:1.刺激T细胞生长。IL-2生物学功能很广泛,能够对多种细胞类型如T细胞、B细胞、NK细胞、巨噬细胞和少突神经胶质细胞等产生作用,其中最显著的作用是影响T淋巴细胞的生长。各种刺激物活化的T细胞一般不能在体外培养中长期存活,加入IL-2则能其长期持续增殖,因此IL-2曾被命名为T细胞生长因子(T cell growth factor,TCGF)。静止的T细胞表面不表达IL-2R,对IL-2没有反应;受丝裂原或其它刺激活化后T细胞才能表达IL-2R,成为IL-2的靶细胞;而IL-2又可诱导靶细胞增加IL-2R的表达。在活体内,IL-2对CD4+T细胞的作用是通过自分泌途径实现的,因为活化的CD4+T细胞能够产生大量的IL-2;而CD8+T细胞则通过旁分泌途径来维持细胞的生长。IL-2R在T细胞上的表达是一过性的,一般在活化后2~3天达到高峰,6~10天左右消失。随着IL-2R的消失,T细胞即失去对IL-2的反应能力。因此若要维持正常T细胞在体外长期生长,必须不断地用丝裂原或其它刺激物去刺激T细胞,以维持IL-2R的表达。2.诱导细胞毒作用。①接受了预刺激信号的CD8+T细胞可以受IL-2的作用活化为CTL,发挥细胞毒作用;在一定条件下,CD4+T细胞也可受IL-2的诱导而具有杀伤作用。②NK细胞是唯一正常情况下表达IL-2R的淋巴样细胞,因此始终对IL-2保持反应性。然而静止的NK细胞上只表达IL-2R的β链和γ链,对IL-2的亲和力低,只能对高浓度的IL-2发生反应。一旦NK细胞活化,就表达IL-2R的α链,成为高亲和力的受体;大剂量的IL-2诱导的LAK活性主要NK细胞。③使T细胞作NK细胞产生IFNγ、TNFβ和TGFβ等因子,促进非特异性细胞毒素;还可诱导产生某些B细胞生长因子以及造血生长因子等,从而发挥相应的生物学作用。3.对B细胞的作用。IL-2对B细胞的生长及分化均有一定的促进作用。活化的或恶变的B细胞表面表达高亲和力IL-2R,但是密度较低;较高密度的IL-2可诱导B细胞生长繁殖,促进抗体分泌,并诱使B细胞由分泌IGM向着分泌IGG2转换。4.对巨噬细胞作用。人类单核-巨噬细胞表面在正常时兴有少量IL-2Rβ链的表达,但是受到IL-2、IFNγ或其它活化因子作用后,可表达高亲和力IL-2R。单核-巨噬细胞受到IL-2的持续作用后,其抗原递呈能力、杀菌力、细胞毒性均明显增强,分泌某些细胞因子的能力也得到加强。
产生:IL-2的产生细胞IL-2主要由T细胞(特别是CD4+T细胞)有受抗原或丝裂原刺激后合成;B细胞、NK细胞及单核-巨噬细胞亦能产生IL-2。
IL-2受体(IL-2R):IL-2的靶细胞包括T细胞、NK细胞、B细胞及单核-巨噬细胞等。这些细胞表面均可表达IL-2R。IL-2R包含3条多肽链:1条为α链,分子量55KD;1条为β链,分子量75KD;另1条为γ链,分子量64KD。α链的胞内区较短,不能向细胞内传递信号,而β链和γ链的胞内区较长,具有传递信号的能力。3种肽链单独与IL-2结合亲和力较低,只有同时表达才能产生高度亲和力。
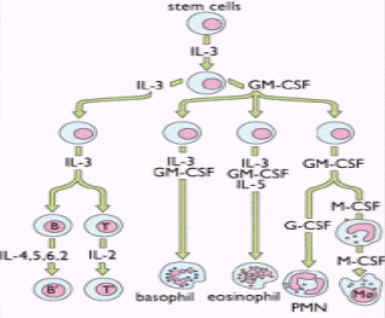
IL-3又称为多能集落刺激因子(Multi-CSF),主要由活化的CD4+T细胞产生。
结构:IL-3分子量约为15KD,其化学本质为糖蛋白。
 白细胞介素3
白细胞介素3 功能:其主要作用为促进骨髓中多能造血干细胞的定向分化与增殖,产生各种类型的血细胞。此外IL-3还可调节多种成熟细胞的生长、分化及相关的基因表达,如C-MYC、IL-2RA基因等。
结构:人和小鼠的IL-4CDna均已克隆成功,并获得了相应的重组产品。成熟IL-4分子量为18~19KD的糖蛋白。
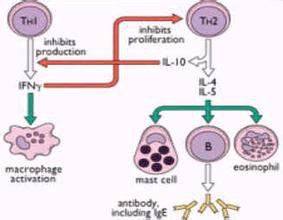
功能:主要由Th2细胞、肥大细胞及嗜碱性粒细胞产生(IL-4由抗原或丝裂原刺激的CD4+T细胞产生,活化的肥大细胞亦可产生IL-4。)。能够促B细胞增殖、分化;诱导IgG1 和IgE产生;促进Th0细胞向Th2细胞分化;抑制Th1细胞活化及分泌细胞因子;协同IL-3刺激肥大细胞增殖等。
IL-4的生物学活性包括以下几个方面:①促使抗原或丝裂原活化的B细胞分裂增殖,但其作用远弱于IL-2;然而IL-4能促使静止的B细胞表达MHCⅡ类分子,增强B细胞的抗原递呈能力,因此曾称为B细胞刺激因子。IL-4是Ig重链基因类转换的主要调节因子,能促使B细胞表达和分泌IgE。此外,IL-4还能诱导B细胞表达低亲和力fc受体。②象IL-2一样,IL-4也是CD4+T细胞的自分泌性生长因子;此外,IL-4还能促进tc细胞的活性。③IL-4不能刺激巨噬细胞增殖,但可增强巨噬细胞的功能;巨噬细胞受刺激后Ⅱ类抗原和fcγr的表达量均明显增加;同时巨噬细胞递呈抗原的能力及对肿瘤细胞的细胞毒素作用也显著增强。④IL-4与IL-3协同可维持和促进肥大细胞的增殖,在某些起敏反应性疾病发生中具有一定的意义;可与CSF协同作用,促进骨髓造血前体细胞的增殖,诱导髓样细胞定向分化;诱导内皮细胞表达血管细胞粘附分子-1(VCAM-1),对淋巴细胞的迁移具有一定意义。
结构:与IL-4一样,IL-5(白细胞介素-5)也由抗原活化的CD4+T细胞产生;肥大细胞也能产生IL-5。天然IL-5的分子量为40~50KD,是由二硫键连接的二聚体糖蛋白,但是单体IL-5也有IL-5活性。
功能:IL-5曾补称为T细胞替代因子(T cell replacing factor,TRF),后来发现人类的IL-5对B细胞没有明显的刺激作用;其主要功能是刺激嗜酸性粒细胞增殖、分化及活化。IL-5不仅使嗜酸性粒细胞的数量增加,而且能增强其功能。在蠕虫感染和过敏性疾病时出现的嗜酸性粒细胞增多主要是由IL-5引起的。人类IL-5还能促进嗜碱性粒细胞释放组胺和白三烯等炎症介质,从而提高嗜碱性粒细胞的活性。
结构:人类IL-6基因位于第7号染色体上;IL-6分子量在21~30KD之间。
功能:主要由单核巨噬细胞、Th2细胞、血管内皮细胞、成纤维细胞产生。能够刺激活化B细胞增殖,分泌抗体;刺激T细胞增殖及CTL活化;刺激肝细胞合成急性期蛋白,参与炎症反应;促进血细胞发育。
IL-6可由多种细胞合成,包括活化的T细胞和B细胞、单核-巨噬细胞、内皮细胞、上皮细胞以及成纤维细胞等。
IL-6作用的靶细胞很多,包括巨噬细胞、肝细胞、静止的T细胞、活化的B细胞和浆细胞等;其生物效应也十分复杂,曾称为B细胞刺激因子2(bsf-2)、26KD蛋白、B细胞分化因子(bCDf)、肝细胞刺激因子(hsf)等。①促进T细胞表面IL-2r的表达,增强IL-1和TNF对TH细胞的致有丝分裂作用。②作为肝细胞刺激因子,在感染或外伤引起的急性炎症反应中诱导急性期反应中诱导急性期反应蛋白的合成,其中以淀粉状蛋白a和c-反应蛋白增加尤为明显。③促进B细胞增殖、分化并产生抗体;多发性骨髓瘤的恶变B细胞既能产生IL-6,又能对IL-6发生应答,提示IL-6可能作为这些细胞的自分泌性生长因子。④IL-6还能有效地促进TNF和IL-1诱导的恶病质;促进糖皮质激素合成;刺激破骨细胞活性和角质细胞生长;还能促进骨髓造血的功能。IL-6不能刺激相应细胞分泌其它细胞因子,在生理浓度下对免疫细胞的自分泌作用亦比较弱,提示其主要免疫学功能是加强其它细胞因子的效果。
结构:IL-7是由骨髓基质细胞分泌的糖蛋白,分子量为25KD;其基因位于第8号染色体。现多采用基因工程手段,从转染含IL-7CDna表达性质粒的哺乳类细胞培养上清中获取IL-7。功能:IL-7的靶细胞主要为淋巴细胞,对来自人或小鼠骨髓的B祖细胞、胸腺细胞及外周成熟的T细胞等均有促生长活性。①与干细胞生长因子(SCF)协同能够刺激b前体发生有丝分裂,这一效应可被tgf和所抑制;但对B祖细胞(pro-b)的生长没有明显作用。②促进双阴性胸腺细胞的成熟,提供胚胎胸腺细胞发育过程中tcr基因重组的始动信号;但对成熟T细胞无明显作用。③诱导胸腺细胞或外周血淋巴细胞产生LAK细胞活性,其效应细胞主要为CD8+亚群;但IL-7诱导的LAK细胞不具有NK活性。④IL-7能刺激髓样前体细胞和巨核细胞产生集落形成单位和血小板,使机体从环磷酰胺的免疫抑制作用中恢复过来;在较高浓度时,IL-7还能增强巨噬细胞的细胞毒活性,诱导单核细胞分泌细胞因子。

结构:主要由单核-巨噬细胞产生;其他如成纤维细胞、上皮细胞、内皮细胞、肝细胞等亦可在适宜的刺激条件下产生IL-8。IL-8的分子量约8KD,主要活性形式为72个氨基酸。IL-8的氨基酸顺序与许多炎症因子有较高的同源性,属于同一个家族;现初步证实IL-8家族(亦称pf4家族)至少有12个成员。IL-8可以分成α和β亚群,α亚群的基因位于第4号染色体上,β亚群的基因位于第17号染色体上。IL-8的受体也有两型:一种只能与IL-8结合,而另一种还能结合其它的趋化因子;中性粒细胞和嗜碱性粒细胞表面均表达丰富的IL-8受体。
功能:主要的生物学活性是吸引和激活中性粒细胞,曾被命名为中性粒细胞激活肽(NAF)、粒细胞趋化肽(GCP)、中性粒细胞激活因子(NAF)等。中性粒细胞与IL-8接触后发生形态变化,定向游走到反应部位并释放一系列活性产物;这些作用可导致机体局部的炎症反应,达到杀菌和细胞损伤的目的。此外IL-8对嗜酸性粒细胞、嗜碱性粒细胞和淋巴细胞也有一定作用。
结构:IL-9主要是由TH细胞产生,成熟分子含126个氨基酸,包括10个半胱氨酸;由于糖基化不同,分子量为30~40KD;经过n-聚糖酶处理后,分子量降到15KD。2-巯基乙醇可使IL-9失活,表明分子间二硫键在维持分子生物学活性方面十分重要。
功能:IL-9可在无IL-2和IL-4的情况下维持TH细胞长期生长;IL-9还可促进某些骨髓样白血病细胞系(如MOTE)的生长,提示其可能还参与造血过程的调控。IL-9与其它白细胞介素的关系尚不清楚,但与IL-2明显的不同是不能诱导CTL和LAK等细胞毒活性,但能维持非抗原依赖性TH细胞的长期生长。此外,IL-9还促进肥大细胞的生长及活性。
结构:IL-10的分子量为35~40KD,通常为二聚体;主要由TH2细胞产生,也可由单核细胞、角质细胞及活化的B细胞产生。
 白细胞介素10
白细胞介素10 功能:主要Th2细胞和单核巨噬细胞产生。能够抑制前炎症细胞因子产生;抑制MHC -II类分子和B-7分子的表达;抑制T细胞合成IL-2、IFN-等细胞因子;可促进B细胞分化增殖。
IL-10能够抑制活化的T细胞产生细胞因子,因此曾称为细胞因子合成抑制因子(csif),特别是抑制TH1细胞产生IL-2、IFN-γ和lt等细胞因子,从而抑制细胞免疫应答。IL-10可降低单核-巨噬细胞表面MHCⅡ类分子的表达水平,损害了APC的抗原递呈能力,实际上这可能是其抑制细胞介导免疫的原因。此外,IL-10还能抑制NK细胞活性,干扰NK细胞和巨噬细胞产生细胞因子;但可刺激B细胞分化增殖,促进抗体生成。
结构:IL-11由骨髓基质细胞产生,分子量约为23KD,是造血微环境中一个多功能的调节因子。
功能:IL-11可刺激浆细胞增殖及T细胞依赖的B细胞发育;促进巨核细胞的形成及成熟,提高外周血血小板数目;与IL-3和IL-4协同作用刺激休止期造血干细胞的增殖;影响红细胞的生成及分化;调节肝细胞血浆蛋白基因的表达,诱导急性期蛋白生成。
结构:IL-12主要由B细胞和巨噬细胞产生;其分子是一种异型二聚体,40KD(p40)和35KD(p35)的2个亚基通过二硫键相连接。
功能:IL-12主要作用于T细胞和NK细胞,曾被命名为细胞毒性淋巴细胞成熟因子(CLMF)和NK细胞刺激因子(NKsf)。IL-12可刺激活化型T细胞增殖,促进TH0细胞细胞向TH1细胞分化;诱导ctl和NK细胞的细胞毒活性并促进其分泌IFN-γ、TNF-α、GM-CSF等细胞因子;促进NK细胞和IL-2rα、TNF受体及CD56分子的表达,增强对肿瘤细胞的adcc效应。
结构:IL-13由TH2细胞产生,分子量约10KD。

功能:IL-13可诱导单核细胞分化,增强其MHCⅡ类分子的表达;抑制lps诱导的单核因子分泌,控制炎症反应;诱导B细胞增殖及合成IgE类抗体,增强B细胞表面MHCⅡ类分子、CD23及CD72的表达;协同IL-2刺激NK细胞产生IFN,从而促进单核-巨噬细胞活化和TH1型细胞免疫反应。最近还发现,IL-13还具有抑制HIV-1在巨噬细胞内复制,诱导中性粒细胞中IL-1RA基因表达和蛋白质合成等多种功能。
结构:IL-14由T细胞分泌,其成熟形式含468个氨基酸残基。
功能:可刺激活化的B细胞增殖,抑制丝裂原诱导的B细胞分泌免疫球蛋白。
功能:IL-15是新近发现的一种因子,可由活化的单核-巨噬细胞、表皮细胞和成纤维细胞等多种细胞产生。IL-15的分子结构与IL-2有许多相似之外,因此可以利用IL-2受体的β链和γ链与靶细胞结合,发挥类似IL-2的生物学活性。属于IL-2家族成员,通过改变Bcl-1家族成员(如Bcl-2和Bcl-XL)的表达来调节T和NK细胞的激活和增殖。
可诱导B细胞增殖和分化,是唯一能部分取代IL-2诱导初期抗体产生的细胞因子;IL-15能够刺激T细胞和NK细胞增殖,诱导LAK细胞活性,还能与IL-12协同刺激NK细胞产生IFN-γ。
白细胞介素-16是由激活的细胞分泌的一种细胞因子。首先被发现可诱导人和大鼠CD4T淋巴细胞的游走应答,因此最初被命名为淋巴细胞趋化因子(LCF)。
结构:人白细胞介素-16由第15号染色体上基因15q26.1编码,其mRNA长2.6kb,半衰期约2h,编码分子质量67*10^3~80*10^3的前体。
功能:近年来研究发现其存在于许多细胞中,对T细胞、肥大细胞、噬酸粒细胞呼吸道上皮细胞等的生长增殖分化、细胞的功能/细胞间信息传递等都具有重要调节作用。
功能:IL-17是Th17细胞主要效应因子,由CD4+ T细胞分泌,能够诱导上皮细胞、内皮细胞、成纤维细胞合成分泌IL-6、IL-8、G-CSF、PGE2,促进ICAM-1的表达。
研究发现IL-17 是T细胞来源细胞因子,该家族包括6个成员的配体(IL-17A~IL-17F)和5个受体(IL-17RA~IL-17RD和SEF)。IL-17是一种主要由活化的T细胞产生的致炎细胞因子,可以促进T细胞的激活和刺激上皮细胞、内皮细胞、成纤维细胞产生多种细胞因子如IL-6、IL-8、粒细胞-巨噬细胞刺激因子(GM-CSF)和化学增活素及细胞黏附分子1(cellular adhesion molecule 1,CAM-1),从而导致炎症的产生。
IL-17是T细胞诱导的炎症反应的早期启动因子,可以通过促进释放前炎性细胞因子来放大炎症反应。IL-17与受体结合后,可通过MAP 激酶途径和核转录因子kB(nuclearfactor kB,NF-kB)途径发挥其生物学作用。Th17细胞能够分泌产生IL-17A、IL-17F、IL-6以及a肿瘤坏死因子(tumor necrosis factora, TNF-a)等,这些细胞因子可以集体动员、募集及活化中性粒细胞。Th17细胞产生的IL-17能有效地介导中性粒细胞动员的兴奋过程,从而有效地介导了组织的炎症反应。
结构:属于IL-1配体家族,结构与IL-1蛋白家族相似。人IL-18cDNA编码193个氨基酸,半胱氨酸天冬酶在N端将其水解为成熟的IL-18,发挥其生物学活性。其位于染色体11q22.2-22.3,由6个外显子和5个内含子组成,cDNA全长约1.1kb。
功能:是一种作用强大的前炎症细胞因子,最具特征的功能是调节细胞增生,分化及细胞外基质生成,故IL-18在糖尿病肾病(DN)的发生、发展中起重要作用。
它是一个前炎症因子,可调节多种细胞发育及细胞因子分泌。研究表明,它是独特的依赖细胞因子周围环境而刺激Thl和Th2细胞反应的细胞因子。能促进外周单个核细胞产生IFN-γ、IL-2和粒细胞巨噬细胞集落刺激因子 等细胞因子,增强NK细胞和Thl细胞的细胞毒作用,促进T细胞的增殖,并在Thl细胞分化和免疫反应中有促进和调节作用。在免疫调节、抗感染、抗肿瘤及慢性炎症性疾病发病过程中起着重要作用。
结构:成熟区153个氨基酸,与IL-10有同源性;染色体1q32。
功能:对抗原呈递细胞具有调节和促增殖效应。活化Stat3,受体为IL20R1/IL20R2。
结构:成熟区164个氨基酸,与IL-10有同源性;染色体1q32
功能:结合IL-20R1/IL-20R2,重组IL-20小鼠腹腔注射可明显刺激中性粒细胞的移动;参与上皮细胞发育,活化角质细胞Stat3,与牛皮癣有关。
结构:成熟区131个氨基酸,与IL-2、IL-4、IL-15空间结构同源,受体包括IL2Rγ链; 染色体4q26-q27。
功能:促进骨髓NK细胞的增殖与分化,与抗CD40抗体协同刺激B细胞的增殖,与抗CD3抗体协同刺激T细胞的增殖。
结构:成熟区146个氨基酸,与IL-10有同源性;染色体12q15。
功能:活化多种细胞系的STAT1,3,包括TP-10(肾癌细胞系)和SW480(肠癌细胞系)。促进炎症时的急性期蛋白产生;结合IL22R/IL10R2 OR IL22BP。
结构:与IL-12有同源性,异源双聚体,α链为p19,含189个氨基酸,与IL-12 p35 同源性;染色体12q13;其β链为IL-12的p40。
功能:经Stat4活化PHA刺激的T细胞,促进其增殖和γ干扰素产生,并诱导记忆性T细胞的增殖。
结构:与IL-10有同源性,206个氨基酸;染色体1q32。
功能:结合IL-22R1/IL-20R2 or IL-20R1/IL-20R2,活化Stat3信号转导途径,促进肿瘤细胞凋亡。
白细胞介素17E(Interleukin 17E,IL-17E),又称作IL-25。
1、结构:与IL-17有同源性, 161个氨基酸;染色体14q11.2 。
2、功能:TH2细胞产生,刺激TH2细胞功能,参与速发型变态反应;支持淋巴样细胞增殖,刺激FDCP2的增殖。 它是由活化的记忆T细胞产生的一种前炎细胞因子,可诱导多种趋化因子和细胞因子的表达,在炎症和造血过程中发挥重要作用。
3、染色体定位:染色体定位分析提示IL-17E位于14q11.2,由2个外显子和1个内含子组成,与CKLFSF5(趋化素样因子超家族成员5,Chemokine-like factor superfamily member 5)紧密连锁,仅相距约500bp。CKLFSF5是我室在国际上首先克隆的趋化素样因子超家族(Chemokine-like factor superfamily,CKLFSF)成员之一。CKLFSF在人类共有9个成员,已有的功能研究提示,CKLFSF代表一个从趋化因子到四次跨膜蛋白之间的过渡性蛋白家族。IL-17E与CKLFSF5的超近距离连锁,提示它们之间可能有一定的相关性。
4、表达谱:人IL-17E是通过同源性分析、序列拼接获得的,到目前为止,尚未用Northern Blot方法在任何组织中检测到其mRNA的存在。IL-17E确切的生理性细胞来源亦尚属未知。用GeneAtalas分析IL-17E的表达谱,发现在舌、支气管上皮细胞和骨髓中,IL-17E表达水平很高;在肾上腺、脑桥、脊髓、胎脑、甲状腺、肾脏、前列腺、肝脏中有中等程度的表达(见图1)。
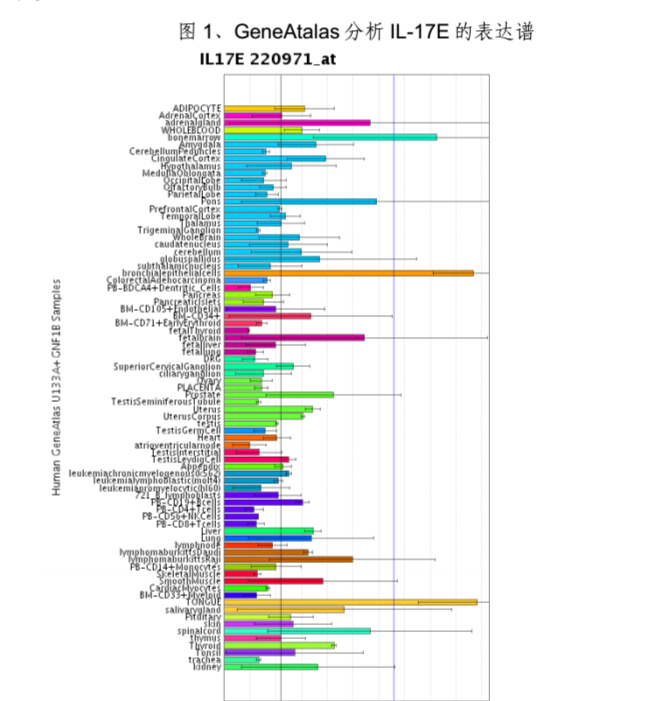 IL-25表达谱
IL-25表达谱 应用RT-PCR技术,在脑、肾脏、肺、前列腺、睾丸、脊髓、肾上腺、气管等多种组织中均可检测到较低水平IL-17E的转录。Ikeda等用RT-PCR和免疫印迹的方法证明,肥大细胞可在IgE作用后产生IL-17E。免疫系统中IL-17E的表达调控仍属未知。
5、蛋白结构:目前发现IL-17E有两种cDNA,一种编码177个氨基酸,另一种编码161个氨基酸,两种蛋白产物羧基端的159个氨基酸残基完全相同,区别仅在于氨基端。信号肽预测和实验证明,距羧基端145个氨基酸残基处为切割位点,两种不同的编码产物分别切割掉了氨基端的33个和16个氨基酸残基,生成相同的由145个氨基酸残基组成的单体。成熟的IL-17E是一种分泌性糖蛋白,为同源二聚体分子,每个单体上有一个N-糖基化位点。
 IL-17家族同源性分析
IL-17家族同源性分析 6、受体:IL-17Rh1(IL-17 receptor homolog 1)是IL-17E的受体。IL-17Rh1也是通过同源性分析、序列拼接获得的,定位于3p21.1,编码502个氨基酸。IL-17Rh1是I型跨膜分子,是IL-17R家族成员,与IL-17R氨基酸水平的同源性为26%,Northern Blot分析提示其在肝脏和肾脏高表达,在脑、结肠、小肠、骨骼肌亦有表达。IL-17Rh1可结合IL-17E和IL-17B,但与前者的亲和力远远高于后者。IL-17E与IL-17Rh1结合后,可活化NF-kB,诱导IL-8等前炎细胞因子的表达,但具体的信号传导通路尚未明确。
7、主要活性:IL-17E的生理功能目前仍不明确,推测其可能与炎症反应的精确调控有关。研究提示,IL-17E可刺激多种来源的细胞系表达IL-6、IL-8等前炎因子和G-CSF(Granulocyte colony stimulating factor,粒细胞集落刺激因子)等造血因子,这一特点与IL-17A十分相似,但目前已知的IL-17E的靶细胞谱比后者窄。
IL-17E转基因小鼠出现严重的多器官慢性炎症,包括肝脏、肺部、心脏、肾脏、淋巴结、脾脏、膀胱。组织器官中有大量中性粒细胞、嗜酸性粒细胞、淋巴细胞、浆细胞和巨噬细胞等多种免疫细胞浸润。小鼠还出现高中性粒细胞和高嗜酸性粒细胞血症,可能与G-CSF和IL-5升高有关。大部分转基因小鼠在6周龄时出现黄疸、血清胆红素和肝酶类升高等肝损表现,有多种细胞因子水平升高,特别是与Th2细胞相关的因子,包括IL-4、IL-5、IL-10、IL-13等,与此一致的是,转基因小鼠出现血清IgE和IgG1的明显升高;但与Th2细胞关联不大的一些细胞因子,如IFN-g(Interferon-g,g-干扰素)、TNF-a(Tumor necrosis factor-a,a-肿瘤坏死因子)、IL-1b等表达水平也有升高。因此,转基因小鼠体内的炎症反应并不仅由Th2细胞介导。另外,IL-17E可抑制关节软骨基质的合成,并刺激一氧化氮的释放和IL-6的产生,提示其可能参与退行性骨关节病的发病。
综上所述,目前的实验结果提示,IL-17E具有多效性,主要参与Th2细胞介导的免疫应答,还与某些前炎因子引起的组织特异免疫病理改变有关。IL-17E可引起并调控多种炎症反应,这提示它有可能在慢性炎症和自身免疫性疾病中发挥作用。
结构:与IL-10有同源性,全长171个氨基酸;染色体12q15。
功能:T细胞产生,可能参与T细胞抗病毒作用。
结构:与IL-12有同源性,异源双聚体,α链为p28,与IL-12 p35同源;其β链为EBI3。
来源:IL-27由p28(IL-12的p35相关多肽)和EBI3(EB病毒诱导的基因3)组成异二聚体。其中p28与IL-12的p35亚基同源,EBI3与IL-12的p40亚基同源。人p28和EBI3基因分别定位于16p11和16p13.3。p28必须与EBI3结合,才具有IL-27生物学活性。
功能:由抗原呈递细胞活化早期阶段产生,促进naive T细胞增殖,与IL-12协同刺激T细胞的γ干扰素产生,促进早期Th1细胞。在T细胞增殖分化早期,可以促进CD4T细胞向Thl方向分化,同时IL-27可以协同IL-2促进初始T细胞产生IFN-γ;促进单核细胞产生IL-1、12、18和IFN等炎症细胞因子。它还可以抑制辅助性17T细胞和诱导型调节性T细胞的发育。研究还发现IL-27可以诱导小鼠B细胞产生LgG2a,抑制IL-4诱导的IgG1的合成;但IL-27可以诱导人B细胞产生LgG1.
归属:属于IL-12/IL-6家族成员。主要由抗原呈递细胞产生,能促进T细胞增殖并向T辅助淋巴细胞(Thl)细胞方向分化,分泌γ干扰素IFN,在抗感染免疫及抗肿瘤免疫等方面发挥作用,参与机体多种疾病的演变过程。
结构:与干扰素及IL-10有低水平同源性,IL-28A与IL-28B有96%同源性;染色体定位19q13.13。IL-28A 200aa,IL-28B 198aa。
功能:抗病毒效应。
白细胞介素-28(IL-28A、IL-28B)和白细胞介素29(IL-29)是一组由病毒或双链RNA诱导的多种细胞产生的新型白细胞介素 ,它们能结合一种由IL-10Rβ和IL-28Rα组成的异二聚体型Ⅱ类细胞因子受体,通过Jak STAT信号通路而发挥其抗病毒或其他防御功能。
人IL-28A、IL-28B和IL-29是由外周血单个核细胞(PMBC)和树突状细胞(DC)等产生的细胞因子,它们组成一个新的细胞因子家族。其基因结构与IL-10相近似,但氨基酸水平与干扰素(IFN)更接近,故Kotenko等也将它们相应地称为IFN-λ1(IL-29)、IFN-λ2(IL-28A)和IFN-λ3(IL-28B)。其基因均位于人19号染色体(19q13.13)。IL-29与IL-28氨基酸的同源性为81%;IL-28A与IL-28B为96%。IL-28A、IL-28B和IL-29与其相应的受体结合而发挥抗病毒感染的作用。
结构:与干扰素及IL-10有同源性,200aa;IL-28A与IL-29有81%的同源性;基因定位19q13.13。
功能:抗病毒效应。
结构:成熟IL-31分子由141个氨基酸组成,含4个α螺旋结构;基因定位于12q24.31。
功能:Th2细胞表达的细胞因子,活化多种Stat分子,参与变态反应和炎症性疾病。
结构:定位于16 p13.3,有4种不同剪切体形式,分别命名为IL-32α,β ,δ,γ。
功能:诱导TNFα和MIP-2的表达; 活化NFκB;诱导p38MAPK的磷酸化。
 白细胞介素32
白细胞介素32 又称IL-1F11。
结构:定位于16 p13.3,有4种不同剪切体形式,分别命名为IL-32α,β ,δ,γ 。
功能:结合IL-1受体家族成员ST2,活化NF-κB和MAPK,促进产生Th2细胞因子,参与变态反应。
结构:该蛋白由241个氨基酸残基组成。
功能:IL-34是2008年首次报道的一种新型白细胞介素,目前已知IL-34具有增强单核细胞活力,调节髓样细胞生长分化,加速破骨细胞形成等功能。
结构:IL-35是最新发现的IL-12细胞因子家族成员,由两个亚基组成的异二聚体,即EBI3和IL-12p35.EBI3由EB病毒感染的B淋巴细胞诱导产生,是编码为34 kDa的糖蛋白,27%的氨基酸与IL-12p40相同;IL-12p35与其他单链细胞因子如IL-6同源.1997年Devergne等发现EBI3和IL-12p35可以聚合为新的造血因子,且在体内形成异二聚体,但未对其功能进行研究.
功能:目前认为Treg细胞及其分泌的细胞因子具有持久的免疫耐受效应,从而推断IL-35在病毒感染的免疫耐受期发挥重要的作用。
结构:除IL-18和IL-33以外,全部人类IL-1家族位于2号染色体的IL-1家族基因簇上。IL-IF7包括12个β链,与IL-1家族成员尤其是IL-8拥有共同的结构模型。存在5种剪切变异体(IL-1F7a-e);其中IL-1F7b(NM014439)是最大的,包括6个外显子中的5个。此外,IL-1FTb可以形成同型二聚体。IL-1F7是目前唯一没有发现鼠同系物的IL-1家族成员。通过蛋白酶-1剪切处理,重组IL-1F7b前体可以分裂为成熟的IL-1F7b。成熟的IL-1F7b转运到细胞核中。这个过程需要蛋白酶-1的参与。
功能:对于在2000年的模拟研究中首次被发现的IL-1F7,人们仍然知之甚少。IL-1F7是一种天然的免疫应答抑制物,能够抑制多种炎症因子的表达,基于其效应的基本本质,研究者认为IL-1家族的这一成员应该被命名为IL-37。研究IL-37有助于加强人们对炎症性疾病的认识,为炎症性疾病的治疗提供新靶点。
结构:白细胞介素(interleukin, IL)-38是最近发现的IL-1家族细胞因子的第10个成员。IL-38的特异性受体为IL-36R,它是IL-36的部分受体拮抗剂。
功能:IL-38可抑制Th17细胞产生IL-17和IL-22等炎症介质,也可抑制IL-36γ诱导产生IL-8,从而抑制炎症反应。
不同种属的IL-2的生物学活性呈现下行性,即沿种系谱向上有约束性,向下无约束性。如人和猿的IL-2几乎可以作用于所有哺乳动物的T细胞,但其它哺乳动物的IL一般很少可以作用于人。对IL-2的氨基酸序列分析表明这与其受体结合部位氨基酸序列较保守有关,以及与氨基酸排列顺序和形成的空间构型也有很大关系。另外一个特点是,IL-2具有很强的种属特异性,鸡IL-2(ChIL-2)与哺乳动物IL-2的同源性仅为20~30%。
由人类由第4号染色体上的一个基因编码。
人类的IL-3基因位于第5号染色体长臂区。
人IL-4基因与IL-3、IL-5一样,位于第5号染色体上。
人类IL-6基因位于第7号染色体上;IL-6分子量在21~30KD之间,其差异是由于肽链的糖基化和磷酸程度不同所致。IL-6由2条糖蛋白链组成;1条为α链,分子量80KD;另1条为β链,分子量130KD。α链缺少胞内区,只能以低亲合性与IL-6结合,所形成的复合物迅即与高亲和性的β链结合,通过β链向细胞内传递信息。
其基因位于第5号染色体上,与IL-4基因紧密连接。IL-13分子的氨基酸顺序与IL-4有20%~25%的同源性,在功能上也与IL-4有许多相似之处。
它与其他具有淋巴细胞趋化活性的蛋白无同源性。
Interleukin-21 (IL-21)是近年来发现的一类新细胞因子,与 IL-2 和 IL-15 具有同源性,含有 4 个IL-2 家族特征的 α 螺旋结构,属 IL-2 超家族。
IL-34和其他人体蛋白都没有相似性,也没有明显的共有结构域和共有基序。它在进化过程中受到较多的约束,在氨基酸水平上,它在不同物种间保守性要高于其受体与其竞争相同受体的配体。
IL-38的基因序列与IL-1受体拮抗剂(IL-1Ra)和IL-36Ra的基因有较高的同源性。
白细胞介素-1家族(Interleukin-1 family,IL-1F)的细胞因子与急性、慢性炎症有关,在天然免疫应答中发挥重要的作用。IL-1家族由11个成员组成,即IL-1F1~IL-1F11,其中IL-1F5、IL-1F6,IL-1F8、IL-1F9、IL-1F10和IF11是通过分析序列同源性、蛋白质的三维结构、基因位点和受体结合特征发现并鉴定的细胞因子,2010年上述细胞因子被重新命名为IL-36Ra,IL-36-α,IL-36β,IL-36-γ,IL-38和IL-33。
白细胞介素-17家族(Interleukin 17 family,IL-17家族),是与白细胞介素17(Interleukin 17,IL-17)具有较高同源性、在脊椎动物进化中高度保守的一组蛋白质,目前共有六个成员,IL-17A(原IL-17)、B、C、D、E和F。其中,IL-17B、C、D、E、F的编码基因,是在人类基因组大规模测序过程中,通过同源性分析、EST序列拼接得到的。
IL-17家族成员与已知分子均无明显同源性。IL-17E与IL-17家族其它成员间的同源性为15-20%。人与小鼠的IL-17E的同源性为80%。IL-17家族的六个成员羧基端的70个氨基酸残基同源性较高,结构上都有五个较保守和另外两个相对变化较大的半胱氨酸残基(见图3所示),这种结构与半胱氨酸结基序(cystine knot motif)极其相似[8]。胱氨酸结基序是在神经生长因子(nerve growth factor,NGF)、转化生长因子β(transforming growth factor β,TGF-β)、血小板来源的生长因子BB(platelet-derived growth factor-BB,PDGF-BB)等生长因子中发现的一种由六个胱氨酸残基形成的结构,在进化中比较保守,具有这种结构的生长因子组成了半胱氨酸结超家族(cystineknotsuperfamily)。
1、临床检验上作为免疫系统状况的指示。
例:全身性感染患者中IL-1ra RN2等位基因和RN2/2基因型的分布频率与正常对照相比明显增加(0.34 vs 0.23, P<0.01;0.12 vs 0.05, P<0.05).IL-1α VNTR及IL-1β AvaI基因多态性(等位基因频率及基因型分布)在两组间均无显著差异.另外,全身性感染患者中基因型A2/2,B2/2和RN2/2病死率明显增加(分别为80%,81%和71%).如果患者同时具有A2,B2和RN2三种等位基因中的两种,则全身性感染更为严重(APACHE II和MODS评分更高),病死率为55%~65%.相反,基因型为A1/1,B1/1或RN1/1的全身性感染患者病死率较低(0~13%).结论与IL-1A或IL-1B基因多态性不同,等位基因IL-1RN2的基因多态性对全身性感染具有易感性.等位基因A2,B2和RN2可能是全身性感染的高危基因标记物。
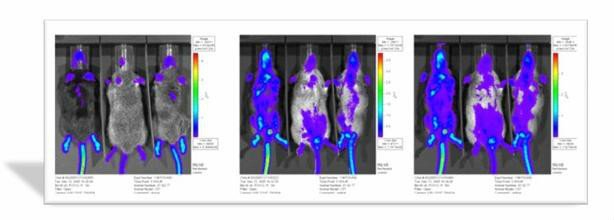 白细胞介素1β光素酶基因表达的转基因小鼠
白细胞介素1β光素酶基因表达的转基因小鼠 2、作为细胞信号调节免疫系统功能,治疗免疫疾病或抗肿瘤。
3、加深对人体生理功能的了解,加深对病情发生、发展的认识。
4、对生命过程的起源进行更深入的认识。
IL-1虽未广泛用于人体研究,但其对免疫系统的特殊作用使它在临床应用方面具有诱人的前景。另外,由于IL-1也参与机体的多种病理过程,所以研究IL-1抑制物可能也具有实际意义。现已克隆出IL-1受体拮抗剂(IL-1 receptor antagonist,IL-1RA),该物质可在体内外封闭IL-1的活性,是一种较理想的IL-1封闭因子。
由于IL-12在抗肿瘤免疫及抗感染免疫中的重要作用,人们对其临床应用寄予厚望;特别是IL-12可协同IL-2促进ctl和LAK细胞的产生表明,IL-12与IL-2联用可望构成一种更有效的肿瘤免疫治疗方法。
由于IL-2能诱导和增强细胞毒活性,目前应用IL-2治疗某些疾病、特别是对肿瘤治疗的研究得到了广泛开展,单独使用IL-2或与LAK细胞等(详见第四章)联合使用治疗肿瘤取得了一定的疗效;还可望用于病毒感染、免疫缺陷病及自身免疫病的治疗。
但IL-2的副作用也日益引起人们的注意:IL-2可引起发热、呕吐等一般症状,还可导致水盐代谢紊乱和肾、肝、心、肺等功能异常;最常见、最严重的是毛细血管渗漏综合征,使患者不得不中止治疗。IL-2的副作用常与IL-2的剂量及用药时间呈相关,停止用药后症状多迅速减轻或消失。IL-2引起副作用的机制是多方面的,但主要是间接性的,即IL-2诱导产生的某些因子或杀伤性细胞起着重要作用;现已知LAK细胞可通过溶解血管内组织而导致多种副作用。给予适当药物(如吲哚美辛、哌替啶、对乙酰基氨酚等)、采取联合用药、改进给药方式(如少量多次短时间输注)和给药途径(如改全身用药为肿瘤局部用药)等将有效地减轻不良反应。
可以用来治疗小儿骨髓增生异常综合征、白血病。
由于IL-3对早期阶段造血细胞的作用较广,可望用于放疗或化疗后患者的骨髓重键。
 关注微信
关注微信