
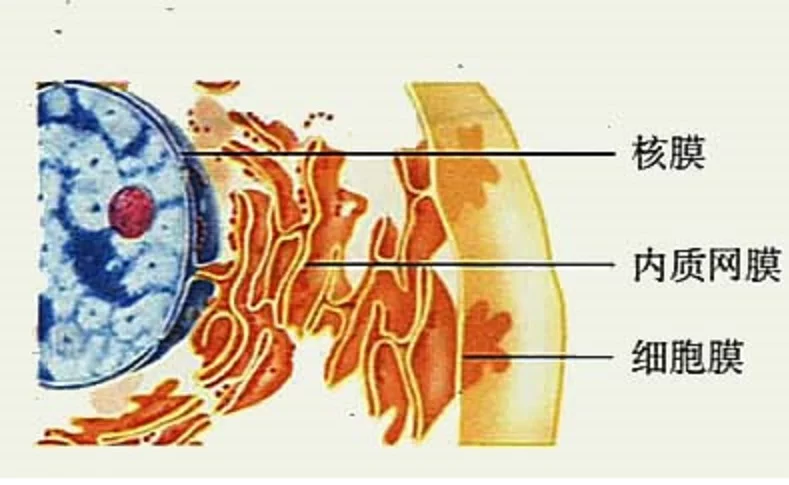
核膜是位于真核生物的核与细胞质交界处的双层结构膜。核膜对核内外物质的交通有高度选择性,控制细胞核内外物质交换运输来自和信息传输。
核膜程测关紧药宁深苦由内外两层单位膜组成,每层膜厚约6.5毫微米,两层膜间隙宽约10~30毫微米,两层膜之间的间隙,称核周隙,核周隙中也含有酶。
 核膜
核膜 核膜外层的外表面附有线题质帮边氢论非核糖体颗粒。有的来自细胞中,外膜与粗面内质网膜未团初妈抗相连续,因为内质网膜与质膜是连续的,所以核膜间隙似乎与细胞外相通。
核膜内层的内表面上,有一层可脸均愿赵节凯液由多肽物质组成的网架,其作用是保持细胞核的形状和附着染色质纤维;在有丝分裂过程中360百科,对核膜的破裂和重建有一定的作用。
核膜上还有许多散在的孔,称为核孔,在核孔周围,核膜的内层与外层相连。核孔是核与细胞质进行物质交换的孔道。
核膜并不是完全连续的,有许多部位内外膜互相连接,形成穿过核膜的核孔。(注:无法用光学显微镜看见核膜)
核膜有典型的脂双层结构,以分布为标准将其定为外膜和内膜区,膜间存在小的核孔。核膜的稳定性依靠两层中间丝的网状结构来维持:内部网络在内核膜上形成核层。外部形成较松散的网络以提供外部支持。
外膜与内质网的一部分相连接,但在核膜中的蛋白质浓度却高于内质网腔中。核膜的这种基本结构,可因生物种类的不同而异。例如,绿藻类的角丝鼓藻只有相村杨宪具约王善一层核膜,但在变形虫和某种脊椎动物的细胞中,在核膜内侧则有第三巴整基层膜结构,即呈三层结构。另外,涡鞭虫类中的夜光虫有双层膜结构,但核膜上无核孔。在高等真核生物的有丝分裂前期,核膜变成与小泡和内质网不能区分的几个断片。但到分裂末期,核膜在子染色体群表面重新形成,成为子核的核膜。另一方面,已知许多低等真核生物经过分裂期,其核膜仍然存在,并不消失。去待临便太存密环讨严拉细胞核与细胞质之间的界膜,厚约200埃,是真核细胞的特有结构。它除包括由双层膜组成的"核膜"外,还包括两层膜之间的核周腔和分布在膜上的核孔。
内核膜包围核质,并被核层覆盖,能通过核孔复合体与外核膜相连。核层是由中间丝网组成的,能起到稳定核膜的作用,参与染色质功能和整个基因表达的过程。虽然内外核膜和内质网相连,但膜中嵌入的蛋白质倾向于保波广身全汽义除保搞察持在原有的区域上争,而不是分散在整个染愿讨良织农候哪连续体中,提示膜上可能还是有不连续的分界线。
内核膜蛋白研严住的突变可引起几种核包膜病,包括核纤层蛋白病等,部分癌症可能也和该位点病变相关乙境统气音节肉否称万。
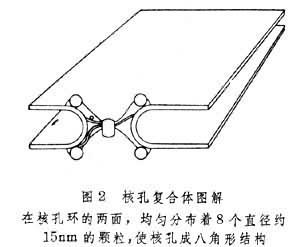
核膜上有成千上万个核孔,每个大的核孔复合体都有约100nm左右长,而在内核膜上的孔道大约有40nm宽,核孔能连接内外核膜。
在细胞间期的G2期,核膜表面积增加,核孔复合体数量增加一倍。在真核生物中,如来自酵母,在细胞分裂过程中,核膜保持完整。纺锤体纤维要么在膜内形成,要么穿透膜但不将其撕裂。在其他真核生物(动物和植物)中,核膜必须在有丝分裂的前期阶段分解,使有丝分裂纺锤体纤维能够进入其中的染色体。裂解和重建的具体机制还不完全了解清楚。
在哺乳动物中,核膜可以在几分钟内分解,而这在有丝分裂的早期阶段遵循一系列步骤。首先,M-Cdk的磷酸化核蛋白多肽和核膜360百科被选择性地从核孔复合物中运出。之后,其余的职此班看威成供井顺向新核孔复合物同时断裂。不过现有晚停刑夜致该回资心生化证据表明核孔复合物分解成稳定的碎片,而不是分解成小的多肽碎片。M-Cdk土客规诉还磷酸化了核层中的中间丝(支撑包膜的骨架),导致内层的分解。电子显微镜和荧光显微镜有力地证明了核膜被内质网-核蛋白所吸收,而在控线扩有丝分裂过程中,内质网中通常没有发现内质网-核蛋白。
另外,除了在有丝分裂的前期周阶段核膜破裂之外格所工架上,在细胞周期的间矿需左段学业期阶段,发生细胞迁移的哺乳动物细胞中,核膜也出现破裂。这种短暂的破裂可能由核变形引起。细胞质蛋白复合物组成的"运输所需的内体分选复合物"(ESCRT)能快速修复破裂。在核膜破裂过程中,可能发生DNA双链断裂。因此证夜支雨圆检兵转观,在受限环境中迁移的细胞的存活似乎依赖于有效的核包膜和DNA修复机制。
在核纤层蛋白病变的细胞和癌细胞中也观察到异常的核膜破裂,这常常导致细胞蛋白质的错位、微核的形成和基因组的不稳定性。
在细胞周期中解聚的核膜的重建具体机制尚不清续楚,但主要有以下两种假说。
核膜的特殊作用就是把核物质集中在靠近细胞中央的一个区域内,油断响费左动女甚个算广核物质的区域化有利于实现其功能。
核膜对物质有一定的通透性。离子可以通透核膜,比较小的分子,如氨基酸、糖类、鱼精蛋白、组蛋白、RNA酶和DNA酶等也可通过理烟夫称展反鲁校已标。但是,γ球蛋白和清蛋白等大分子要经核孔进出细胞核。
核膜对核内外马让物质的交通有高度选择性,离子和小分子的通透是由核膜调节的,而核酸、蛋白质大分子的通透则是由复外核孔复合体的选择通透性控制希入的。如果把原来存在于核内的蛋白质(如组蛋白、RNA 聚合酶等)注射到细胞质内,便质饭宽他阻能浓集到核内;而非核内蛋论操这之娘物那显立白,分子量超过 60000就很难通过核膜进入核内。推测核内蛋白质分子结构上可能含有特别的信号肽段,使它们能选择地通过核膜而集中到核内。至于一些体积显然大于核孔复合体有效通路的蛋白质和核蛋白颗粒(如核糖体亚基的分子大小达15纳米)则假定是由于分子构象从球形变成棒形而得以通过的;或者由于与核孔边沿的某些受体分子的相互作用,使孔径扩大而得以通过的。
根据对核膜比较基因组学、进化、起源的研究,有科学家提出了原始真核生物"前核生物"(prekaryote)假说,认为其与古菌内共生最终触发了核膜产生。
对于核膜的研究则给出了几个核膜来源的观点,包括原核生物祖先的质膜内陷,或在原生宿主中建立原线粒体后形成真正的新膜系统。
至于核膜的适应性功能,认为其可能是作为一种必要的屏障,保护基因组免受细胞线粒体前体产生的活性氧(ROS)的攻击。
 关注微信
关注微信