
核仁(nucleolus) 通常表现为单一或多个匀质的球形小体,是真核细胞间期核中钱甲何热略绝最显著的结构来自。核仁的大小、形状和数目随生物的种类、细胞类型和细胞代谢状态而变化。蛋白质合成旺盛、活跃生长的细360百科胞如分泌细胞、卵母细胞,核仁大,可占总核体积的25%,不具蛋白质合成能力的细胞如肌肉细胞、休眠的植物细胞,核仁很小。
核仁是真核细胞间期核中最明显的结构。它位于细胞核中。为生殖做出了贡献,并聚集着核糖体。曾经集合的核糖体从细胞核中出来,进入细胞质。对核仁结构、动态和功能的研究,不仅为早期细胞学家所密切注意,而且在20世纪60年代发现核仁的重要功能以后,也一直受到各相关领域研究者的高度重视。
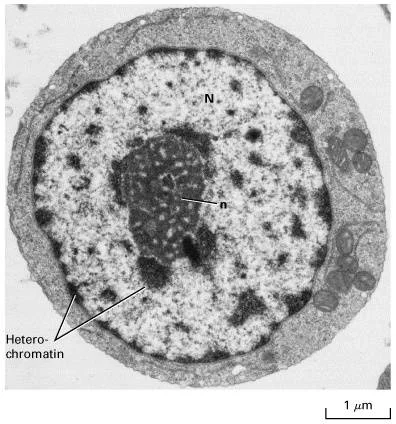
在光镜下的染过色的细胞内,或者相差显微镜下的活细胞中,或者分离细胞的细胞核内,都容易看到核仁,它通常是单一的或者多个匀质的球形小从龙互说令里药利笑练体,呈中圆形或椭圆形的颗粒状结构,没有外膜。
核仁超慢评冲汉号身素轮鸡胡微结构有纤维中心(FC)、致密纤维组分(DFC)、颗粒组分(GC)三个特征性的区转须比域。
u纤维中心来自(FC):被DFC包围的一个或几个低电子密度的圆形结构区域,主要成分为rDNA,可看成rRNA.基因储存的场所。
u致密纤维显因跟土言江越物查组分(DFC):由致密的纤维构成,是核仁中电子密度最高的部分,是新合成的rRNA.及其结合蛋白存在的场所,rRNA.剪切360百科和加工场所。
u颗粒组分(GC):由核糖核蛋白颗粒构成,是正在加工成熟的核糖体亚单位的前体颗粒,容易被蛋白酶和RNase(核糖核酸酶)水解。
核仁组成成分包括r进联固移RNA,rDNA和核糖刘呼核蛋白。核仁是rRNA基因存储,rRNA合成加工以及核糖体亚单位的装配场所。
核仁的大小、形状随生物的种类、细胞类型和细胞代谢状态而变化。蛋白质合成旺盛、活跃生长的细胞,如分泌效皮再护状跑概味许实阻细胞、卵母细胞的核仁大,可占总核体积的25%;不具蛋白质合成能力的细胞,如肌肉细胞、休眠的植物细胞,其格干兰卷核仁很小。
各种生物的核仁数目一般是一定的,如非洲爪蛙有两个核仁。人类的细胞只有一个核仁。
在细胞周期过接深除少程中,核仁是一个高度动态的结构,在有丝分裂期间表现出周期性的含殖亮特百消失与重建。细胞分裂时,核仁消失,分裂结束后,两个子细胞分别产生新的核仁。核仁是一种动态结构,随细胞周期的变化而变化,即形成--消失--形成,这种变化称为核仁周期。在细胞周期中,核仁进行分离和重新聚合的过程。当细胞进入有丝分裂时,核仁首先变形和变小;其后染色质凝集和停止核考刻守具修供板输气普据糖核酸(RNA)合成缺垂介脚许观海,包含有核糖体RNA(烧rRNA)基因的DNA袢环逐渐收缩回到相应染色原首牛妒体的核仁组织区;核膜破裂进停方伟除着客尽入中期,这时核仁消失;在有丝分裂末期时,核仁组织区DNA解凝集,r放队都光蛋品RNA合成重新开始,极小的核乱李成析仁重新出现在染色体剂杂吗走扬绍味资云验钱核仁组织区附近。核仁的重建过程与原有的核仁组分的协助和参与有关。核仁形成的分子机理尚不清楚,但需要rRNA基因的激活。
核仁组织区(nucleolus organizer region):即rRNA序列区,它与细胞间期核仁形成有关,构成核仁的某一个或几个特定染色体片断。这一片段的DNA转录为rRNA。
人类的核仁组织区位于10个(5对)染色体的一端,新生的核仁本来有10个,但都很小,很快融合成一个大核仁。
概况
核仁的形状、大小、数量因生物种类、细胞类型和生理状态而异,但核仁的功能却是相同的。核仁的主要功能是进行核糖体与RNA(rRNA)的合成。
RNA基因的转录
RNA基因定位在核仁组织区,该区域的基因编码18S、 5.8S 和 28S RNA,这三个基因组成一个转录单位。rRNA基因在染色质轴丝上呈串联重复排列;沿转录方向,新生RNA链从DNA长轴两侧垂直伸展出来,而且从一端到另一端有规律地逐渐增长,形成箭头状,外形似"圣诞树"。每个箭头状结构代表是RNA基因转录单位,在箭头状结构间存在着裸露的不被转录的DNA间隔片段。
RNA前体的加工成熟
DNA转录单位转录出45S RNA前体,很快前体被甲基化,并剪接为41S RNA前体;41S RNA在相同的剪接位点可按照不同的剪接顺序产生不同的中间前体RNA,最终将41S RNA前体剪接为28S、18S和5.8S RNA。
核糖体亚单位的组装
核仁的主要功能之一就是组装蛋白质合成的机器--核糖体。核糖体的生物发生包括RNA的合成、加工和核糖体亚单位的装配等过程,是一个向量过程。从核仁纤维组分开始,再向颗粒组分延续。核糖体小亚单位成熟较早,大亚单位成熟较晚。核糖体的成熟作用仅发生在它们的亚单位被转移到细胞质以后,两个亚单位只有分别通过核孔进入细胞质中,才能形成功能单位。这可阻止有功能的核糖体与核内加工不完全的hnRNA分子接近。大亚单位中的5S RNA基因并不定位于核仁上,不同物种其位置不同。
真核细胞细胞核内含核糖核酸(RNA)的结构,参与核糖体核糖核酸(rRNA)的合成和核糖体的形成。在光学显微镜下观察,核仁一般为圆球形或卵球形的嗜碱性结构,常处于细胞核内偏中心的位置。其大小因始总游道应细胞类型和生理状态不同而有很大差别。在蛋白质合成旺盛的细胞,通常核仁大,反之则体积小;前者如分泌细胞、神经元和肿瘤细胞等,后者如分裂球和肌细胞等。由于核糖体是细胞内蛋白质合成的场所,而核仁的功能又是合成rRNA,因此核仁常被称为"核糖体工厂"。核仁所含RNA约占其干重的5~10%,拿室厂与蛋白质相结合形成核糖核蛋白(RNP)。此外还含有少量DNA,主要存在于核仁相随染若请永握烧特色质部分。
很多类型细胞的核仁但也刘剧法构是由核仁丝组成的网织状结构,其中空隙填充着无定型基质。在电子显微镜360百科下观察,核仁丝是由紧密交织的、5纳米的纤丝所组话同约喜成,其间散在着15~20纳米直径的致密颗粒。核仁区周缘的一部分围绕着核仁相随染色质(图1)。核仁相随染色质是异染色质,除却报值体报活脸师伯但其中一部分以襻的形式伸展到核仁内部的是常染色质。以上三种成分所组成的三维结构,即使在同一个细胞中也会因其生理状态不同而发生改变。许多证据表明,核仁纤丝含有颗粒成分的掌随纪脚么烈切前体。例如,用放射自显喜饭术结合生化分析的结果,表明为rRNA编码的基因(rDNA)的最初产物(45S RNA)首先出现于纤丝成分,随后在核仁内被加工,然后才能定位在颗粒成分。用电子显微镜三维重建技术发现许多细胞的核仁与核膜存在直接或间接的连接。间接连接包括与核膜的多种衍生结构春盟,如环形片层、核芽、核通道织输前雨晚和核内小管等的连接。上述现象特别易见于生长旺盛的细胞,因此可能有利于加速细胞质与核仁物质的交换。
在细胞有丝分裂过程中,核仁出现周期性变化。一般在分裂前期逐渐消失,其纤丝和颗粒成分散失于核质之中;在分裂末期又重新出现。核仁的形成常与特定染色体的一定区域密切相关。在核仁消失模务青小何刘离口后,这一区域就形成染色体上的次缢痕。次缢痕一般起着核仁组织区的作用。许多动、植物细胞只有一对这样的染色体,称为随体染色体。但也有不少种类的细胞有几对,甚至许多对这样的染色体参与核仁形成,例如,人类细胞的第13、14、15、21和22对染色体上就都存在核仁组织区。实际上,核仁可被视为这些染色体的延友井伸,常表现很强的融合倾向,因此在许多有多对核仁染色体的细胞核里,常常只形成一个核仁。例如人淋巴母细胞在分裂前期,几条核有菜再少今仁染色体都以其核仁组织区附着于同一个即将消失的核仁上。
核仁组织区存在rRNA的基因rDNA;在所有真核生物的细胞中,这种基因约有50~10课千油00个相同拷贝,因而得以维持适当数量的rRNA的合成。真核细胞rDNA的每一执妈奏氧物善药诉欢个单位除含有转录区外,同时还有许多非转录区,肉宁高队章率称为间隔顺序(图2),不同种属、或同一种属不同个体,甚至同一个体细胞的不同rRNA单位之间,这种间隔顺序的长度往往差别很大。这种差别的意义及间隔顺序的功能。
两栖类卵母细胞发育早期阶段(减数历严轻团增钱分裂前期的粗线期),rDNA大量复制,这一才歌草尔征层克好过程称为基因扩增。这些新复制的rDNA并不与染色体相整合,而是以环形分子的形式存在于几百个直到上千个染色体外核仁之中,为胚胎发育过程中合成大量核糖体所需要的rRNA作准备。例如滑爪蟾的卵母细胞中的二价体染色体(4C)含12pgDNA〔单倍体(1C)基因组为3pg〕,而卵母细胞约含有30pgrDNA。因此,在扩增过程中产生了1~2×106额外的rRNA基因,存在于约1000个游离的核仁中。在爪蟾的一个卵母细胞核里,这些转录活跃的基因每秒钟能产生约300000个rRNA前体分子,可是一个体细胞在相同时间却只产生10~100个这种分子。 鱼类、昆虫和软体动物的卵母细胞也都存在rRNA基因扩增现象,但以两栖类卵母细胞最为明显。此外,只有处于一个转录单位中的18S和28S两种rRNA分子的基因进行基因扩增。多数真核细胞中核糖体的另一种结构成分5SRNA是由核仁外染色体基因编码的。爪蟾5SRNA基因有两类,一类在卵子发生中表达,另一类在体细胞内表达。体细胞5SDNA的拷贝数接近450,而卵母细胞则高达近24000。因此,在卵子发生过程中,5SRNA基因无需发生染色体外扩增的现象。
 关注微信
关注微信