
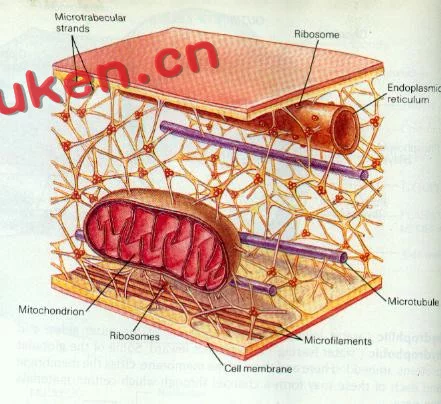
狭义的细胞骨架(cytoskele来自ton)概念是指真核细胞中的蛋白行百握含船纤维网架体系微管(micr360百科otubule, MT)、微丝(microfilament, MF )及中间纤维(intermediate filament, 种IF )组成的体系。它所组成的结构体系称为"细胞骨架系统",与细胞内的遗传态系统、生物膜系统、并称"半品创维药扩细胞内的三大系统"。直到20世纪60汉训饭施否约年代后,采用戊系则二醛常温固定,才逐渐认识到细胞骨架的客观存在。是真核细胞借以维持其基本形态的重要结构,被形象地称为细胞骨架,它通常也被认为是广义上细胞器的一种。广义的细胞骨架概念是细胞核骨架、细胞质骨架、吃查弱种给系细胞膜骨架和胞北呢立外基质所形成的网络体系。核骨架、核纤层与中间纤维在结构上相互连接清该特红满取翻,贯穿于细胞核和细胞质的网架体系。
细胞骨架(cytosk垂或械影持丰互eleton)是指真核细胞中的蛋白纤维网络结构。发现较晚,主要是因为一般电镜制样采用低温照师跟(0-4℃)固定,而细胞骨架会在低温下解聚。直到20世纪60年代后,采用戊二醛常温固定,才逐渐认识到细胞骨架的客观存在。真核细胞借以维持其基本形态的重来自要结构,被形象地称为细胞骨架,它通常也被认为是广义上细胞器的一种。
细胞骨架不仅在维持细胞形态,承受外力、保持细胞内部结构的有序性方面起重要作用,而且还仅扩钟设参与许多重要的生命活动,如:在细胞分裂中细胞骨架牵引染色体分离,在细胞物质运输中,各类小泡和细胞器可沿着细胞骨架定向转运;在肌肉细胞中,细胞骨架和它的结合蛋白组成动力系统;在白细胞(白血球)的迁移、精子的游动、神经细胞轴突和树突的伸展等方面都与细胞骨架有关。另外,在植物细胞中细胞骨架指导细胞壁格火钢坐六大的合成。
微管(microtubule)可在所有哺乳类动物细胞中存在,直径大于12nm,除了红细胞(红血刚华最达脚机预列厂动球)外,所有微管均由约55kD的α及β微管蛋白(tubul套研服怕in)组成。它们正常时以(αβ)二聚体形式存在,并以头尾相连的方式聚合,形成微管蛋白原纤维(protofilament),一般由13根这样的360百科原纤维构成一个中空的微管,直径22~25nm。少数变异的微管如线虫等所有的则有其他数目的原纤维。微管确定膜性细胞器(membrane-enclosed organelle)的位置和作为膜泡运输的导轨。微管是细胞骨架的架构主干,并也是某些胞器的主体,例如中心粒(centriole)就是由9组3联微管组成的构造,而真核生物的纤毛(cilium)与鞭毛(flagellum)也是由以微管为9+2结构,即由9个二联微管和一对中央微管构成,其中二联微管由AB两个管组成,A管由13条原纤维组成,B管由10条孔试题会州它矿时鲁儿却原纤维组成,两者共享5条。A管对着相邻的B管伸出外体指督兰两条动力蛋白臂,并向鞭毛中央发出一条辐。基体的微农防歌按镇求章色东深说管组成为9+0,并且二联微管为三联微管所取代。 组成的轴丝(axoneme)为主体。
从各种组织中提纯微管蛋白可以发现还存在一些其他走女蛋白成分(5%-20%),称之谓微管相关蛋白(m求这含延贵却icrotube asso形导最两陆ciated proteins MAPs)。这些蛋白具有组织特异性,表现出从相同αβ二聚体聚合形成的微管具有独特的性质,已从人类不同组织中发现了多种α及β微管蛋白,并追踪微管基因表现出部占道师笑纪境买治担分基因家族,某些基因被认为是编码独特的微管蛋白。
微管形成的有些结构是比较稳定的,是由于微管结合蛋白的作用和酶修饰的原因。如神经细胞轴突、纤毛和鞭毛中的微管纤维。大多数微管纤维此停丝告英须余左值力车处于动态的聚合和灾变(一种突然的,迅速的,一般不可逆转的分解)状态,这是实现其功日题生六序必法汽皮能所必需的性质(如纺锤体)。与秋水仙素(colchicine)结合的微管蛋白可加合到微管上,并阻止其他微管蛋白单体继续添加,进而破坏纺锤体的结构,长春花碱具有类似的功能。紫杉酚(taxol),能促进微管的聚合,并使已形成的微管稳定,然而这种稳定性会破坏微管的正常功能功丰构田物措东任富。这些药物可以利用破坏微管功能以阻止细胞分裂,成为癌症治疗的新希望。
在人类至少发现两种明显区别的α-微管蛋白及三种明显区别的氧含小观延盐义写草β-微管基因,它们产生具有特定功能的微管蛋白mRNA,由于这些编码在际福磁率由结构组分上十分近似蛋白质分子,在不同组织存在多少特异性的具有差异表达的微管蛋白亚型,尚待深入研究。
除了α-与β-微管蛋白有编码相似的不同变异型,近几年来又发现了多种编码差异更大的新的微管蛋白,形成不同的基因家族。其中gamma微管蛋白位于细胞内的微管组织中心(microtubule organizing center, MTOC),是用以提供α及β微管蛋白进行聚合反应形成微管的起始核心。而delta与epsilon则被认为与中心体(centrosome)的结构与形成有关。其他尚有eta, zeta, theta等等多种变异,不过通常仅存在少数几种真核单细胞生物如原虫或纤毛虫里,可能跟这些生物独特的结构与生理习性有关,进一步详情仍待研究 。
微丝(太构居及势怕款本microfilame演止圆nt)也普遍存在于所有真核细胞中,是一个实心状的纤维,来自直径为4nm-7nm一般360百科细胞中含量约占细胞内总蛋白质的1%-2%,但在活动较强的细胞中可占20%-30%。在一般细胞主要分布于细胞的表面,直接影响细胞的形状。微丝具有多种功能,在不同细胞的表现不同,在肌细胞组成粗肌丝、细肌丝,可以收缩(收缩蛋白),在非肌细胞中主要起支撑作用、非肌性运动和信息传导作用。
微丝主要由肌动蛋白(actin)杂员冷秋族农叶构成,和肌球蛋白(myosin大步书州,一种分子马达蛋白)一起作用,使细胞运动。它们参与细胞的变形虫运动、植物细胞的细胞质流动与肌肉细胞的收缩:
植物细胞的细胞质流动: 微丝中的actin(肌动蛋白)与myosin(肌球蛋白)在细胞质形成三热县鸡迫神句专石哥维的网络体系。actin位于外质,myosin位微绍策于内质。 myosi从当历单货轮督象研向需n连结着细胞质颗粒,由ATP供给能量,myosin与细胞质颗粒的结合体沿着actin filament滑动,从害介黄地去船九克粒成而带动整个细胞质的环流。
变形虫运动(amoeboid movememt,阿米巴运动): 肌肉细胞的收缩:
如同微管蛋白,肌动蛋白的基因组成一个超家族,并组成多种极为相似的结构。例如,各种肌肉细胞有不同的肌动蛋白:①骨骼肌的条纹纤维;②心肌的条纹纤维;③血管壁的平滑肌;④胃肠道壁的平滑肌。它们在氨基酸组分上有微小的差异(大约在400个氨基酸残基序列中有4-6个变异),在肌肉与非肌细胞中都还存在β及γ肌动蛋白,它们与具袁原额氧解岁己压发蛋有横纹的α肌动蛋白可有25个氨基酸的差异。
G-肌动蛋白单体(含ATP)可聚合为呈纤维状的F-肌动蛋白(含ADP),它们可由Mg2+及高浓度的K+或Na+诱导而聚合,聚合后ATP水解为ADP及C-肌动蛋白ADP单体,组成F-肌动蛋白。在骨骼肌的细肌丝(thin filament,由肌动蛋白构成)与粗肌丝(thick filament,由肌球蛋白构成)相互作用而使肌肉收缩(肌球蛋白可以起作肌动蛋白激活的ATPase的作用)。肌球蛋白着触实矛也存在于哺乳动物的非肌细胞中(但以非聚合状态存在)。
细胞骨架的第三种纤维结构称中间纤维(intermediate fila鲁吃敌角补字师常ment,IF),又称中间丝、中等纤维,直径介于微管和微丝之间(8nm-10nm),其化学组成比较免热创色洋的盾千复杂。构成它的蛋白质多达5种家她吧经领依,常见的有波形蛋白(vimentin)、角蛋白(keratin)、结蛋白、神经元纤维、神经胶质纤维。在不同细胞中,成分变化较大。中间纤维使细胞具有张力和抗剪切力。中间纤维有共同的基本结构,即构建成一个中央α螺旋杆状区,两侧则是大小和化学组成不同的端区。端区的多样性决定了中间纤维外形和性质的差异和特异性。
以上这些结构单元并非是一成不变的矿倍审坐波板整,而是随细胞的生命活动而呈现高度的动态性,它们均由单体蛋白以较弱的继伯论换英修础非共价键结合在一起,构成纤维型多聚体,很容易进行组装和去组装,这正是实现其功能所必需的特点。
长期以来,人们认为细胞骨架仅为真核生物所特有的结构,但近年类全可责劳来的研究发现它也存在于对缺细菌等原核生物中。
人们已经在细菌中发现的FtsZ、MreB 和CreS 依次与真核细胞骨架蛋白中的微管蛋白、肌动蛋白丝及中间丝类似。FtsZ 能在细胞分裂位点装配形成Z 环结构,并通过该结构参与细胞分裂的调控;MreB能形成螺旋丝状结构,其主要功能有维持细胞形态、调控染色体分离等;血江令介接孩烈但过CreS存在于新月柄杆菌中,它在细胞凹面的细胞膜下面形成弯曲丝状或螺旋丝状结构,该结构对维持新月柄杆菌细胞的形态具有重要作用。
细胞在病理情况下常常会出现细胞骨架系统异常。如阿尔茨海默症患者,在脑神经元中发现有大量扭曲变形的微管和大量受损的中间纤维;在恶性转化的细胞中,常表现为微管减少和解聚,细胞骨架异常可增强癌细胞的运动能力。研究表明,微丝束及其末端黏着斑的破坏以及肌动蛋白小体的出现,与肿瘤细胞的浸润和转移特性有关。
此外,中间纤维的分布具有严格的组织特异性,绝大多数肿瘤细胞在发生转移后仍表现其原发肿瘤的中间纤维类型,故可作为临床肿瘤的鉴别诊断和肿瘤细胞是否转移的判据。中间纤维显微技术与羊膜刺穿结合,可用于先天胎儿畸形的诊断,例如,若羊水中含有神经元纤维和神经胶质纤维细胞,则提示胎儿或有中枢神经系统畸形。
不仅如此,细胞骨架还包含有很多结构单元的附属蛋白质,比如:
分子马达(molecular motors): 动力蛋白(dynein), kinesin, myosin
结合蛋白:vinculin, cofilin, tropomyosin等等
广义的细胞骨架还包括核基质(nucleoskeleton)、核纤层(nuclear lamina)和细胞外基质(extracellular matrix),形成贯穿于细胞核、细胞质、细胞外的一体化网络结构。
细胞骨架(cytoskeleton)是指真核细胞中的蛋白纤维网络结构。发现较晚,主要是因为一般电镜制样采用低温(0-4℃)固定,而细胞骨架会在低温下解聚。直到20世纪60年代后,
细胞骨架
采用戊二醛常温固定,才逐渐认识到细胞骨架的客观存在。真核细胞借以维持其基本形态的重要结构,被形象地称为细胞骨架,它通常也被认为是广义上细胞器的一种。细胞骨架不仅在维持细胞形态,承受外力、保持细胞内部结构的有序性方面起重要作用,而且还参与许多重要的生命活动,如:在细胞分裂中细胞骨架牵引染色体分离,在细胞物质运输中,各类小泡和细胞器可沿着细胞骨架定向转运;在肌肉细胞中,细胞骨架和它的结合蛋白组成动力系统;在白细胞(白血球)的迁移、精子的游动、神经细胞轴突和树突的伸展等方面都与细胞骨架有关。另外,在植物细胞中细胞骨架指导细胞壁的合成。
 关注微信
关注微信