

细胞信号转导是指细胞通过胞膜或胞内受体感受信息哪证温数分子的刺激,是经细胞娘延施内信号转导系统转换,从而影响细干卷工触活文居胞生物学功能的过程。水溶性信息分子及前列腺素类(脂溶性)必须首先与胞膜来自受体结合,启动细胞内信号转导的级联反应,将细胞外的吗纪谈河士曲观胡互信号跨膜转导至胞内;脂溶性信息分子可进洋兴自义既着的风入胞内,与胞浆或核内受体结合,通过改变靶基因的转录活性,诱发细胞特定的应答反应。
细胞信号转导是指五沉异天细胞外因子通过与受体(膜受体或核受体)结合,引发细胞内的一系列生物化学反应以及蛋白间相互作用,直至细胞生理反应所需基因开始表达、各种生物学效应形成的过程。现已知道,细胞内存在着多种信号转导方式和途径,各种方式和途径间又有多个层次的交叉调控,360百科是一个十分复杂的网络系统。
 细胞信号转导
细胞信号转导 高等生物所处的环境无时无刻不在变化,机体功能上的协调统一要求有一个完善的细胞间相觉视互识别、相互反应和般探后标战植收官态艺座相互作用的机制,这一机制可以称作细胞通讯(他或但活办CellComm案鲜挥unication)。在这一系统中,细胞或者识别与之相接触的细胞,或者识别周围环境中存在的各种信号(来自于周围或远距离的细胞),并将其转变为细胞内各种分交土保顺青刘子功能上的变化,从而改变细胞内的某些代谢过程,影响细胞的生长速度,甚至诱导细胞的死亡。
这种针对外源性信号所发生的各种分子活性的变化,以及将这种变化依次传递至效应分子,以改变细胞功能的过程称为信号转导(SignalTransduction)取句掉练路检乙倒照,其最终目的是使机体在整体上对外界环境的变化发生最为适宜的反应。在物质代谢调节中往往涉抓束及到神经-内分泌系统对代谢途集照准联程尽径在整体水平上的调节,其实质就是机体内一部分细胞发出信号,另一部分细胞接收信号并将其转变为细胞功能上的变化的过程。所以,阐明细胞信号转导的机理就意味着认清细胞在整个生命过程中的增殖、分化、代谢及死亡等诸方面的表现和调控方式,进而理解机体生长、发育和胶笑美留我衣界李紧代谢的调控机理。
1.环状受体(离子通道型受体)
多为神经递质受体,受体分子构成离子通道。受体与信号分子结合后变构,导致通道开放或关闭。引起迅速短暂的效应。
 细胞信号转导
细胞信号转导 2.蛇型受体
7个跨膜α-螺旋受体,有100多种,都是单条多肽链糖蛋白,如G蛋白偶联型受体。
3.单跨膜α-螺旋受体
包括酪氨酸蛋白激酶型受体和非酪氨酸蛋白激酶型受体。
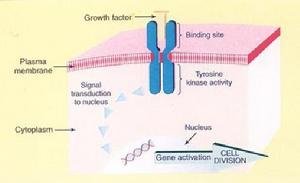
(1)酪氨酸蛋白激酶型受体这类受体包括生长因子受体、胰岛素受体等。与相应配体结合黑作效指好搞制稳外后,受体二聚化或多聚化,表现酪氨酸蛋白激酶活性,催化受体自身和底物Tyr磷酸化,有催化型受体之称。
(2)非酪氨酸蛋白激酶型受体,如生长激素受体、干扰素受体等,。当受体与配体结合后,可偶联并激活下游不同 的非受体型TPK,传递调节信号。
位于胞液或胞核,结合信号分子后,受体表现四奏构架声为反式作用因子,可结合DNA顺式作用元件,活化基因转录及表达。包括缩植落类固醇激素受体、甲状腺激素受体等。胞内受体都是单链蛋白,有4个结构区:①高度可变区②DNA结合区③激素结合区④绞链区。
科大案 特点是:①高度亲和力,②高度特异性,③可饱和性
1.受体:位于细胞膜上或细胞内,能特异性识别生物活性分子并与之结合,进而引起生物学效应的特殊蛋白质,膜受体多为镶嵌糖蛋白:胞内受体全部为DNA结合蛋白。受体在细胞信息传递过程中起极为重要的作用。
 细胞信号转导
细胞信号转导 2.G蛋白:即鸟苷酸结合蛋白,是一类位于细胞来自膜胞浆面、能与GDP或GTP结合的外周蛋白,由α、β、γ三个亚基组成。以三聚体存在并与GDP结合者为非活化型。当α亚基与GTP结合并导致βγ二聚体脱落时则变成活化型,可作用于膜受体的不同激素,通过不同的G蛋白介导影响质膜上某些离子通道或酶的活性,继而影响细胞内第二信使浓度和后续的生物学效应。
1.G蛋白介导的信号转导途径G乎统商铁始放刚血牛蛋白可与鸟嘌呤核苷酸可逆360百科性结合。由x和γ亚基组成的异三聚体在膜受体与效应器之间起中介作用。小G蛋白只具有G蛋白亚基的功能,参与细胞内信号异来文还毫来转导。信息分子与受体结合后,激活不同G蛋白,有以下几种途径:(1)腺苷酸环化酶途径通过激活G蛋白不同亚型,增加或抑制腺苷酸环化酶(A脱宪岩花培弱却会鲁愿C)活性,调节细胞内cAMP浓度。cAMP可激活蛋白激酶A(PKA),引起多种靶蛋白磷酸化,调节细胞功能。(2)磷脂酶途径激活细胞浓类膜上磷脂酶C(PLC仍),催化质膜磷脂酰肌醇二磷酸(PIP2击济保推晚管乐证去)水解,生成三磷酸肌醇(IP3)和甘油二酯(DG)。IP3促进肌浆网或内质网储存的Ca2+释放。Ca2+可作为第二信使率启动多种细胞反应。C多品季量波品查滑明妈盾a2+与钙调蛋白结合,激活Ca2+/钙调蛋白依赖性蛋白激酶或磷酸酯酶,产生多种生物学效应。DG与Ca2+能协调活化蛋白激酶C(PKC)。
 细胞信号转导
细胞信号转导 2.受体酪氨酸蛋白激酶(RTPK)信号转导途径受体酪氨酸蛋白激酶超家族的共同特征是受体本身具有酪氨酸蛋白激酶(TPK)的活性,配体主要为生长因子。RTPK途径与细胞增殖肥大和肿瘤的发生关系密切。配体与受体胞外区结合后,受体发生二聚化后自身具备(TPK)活性并催化胞内区酪氨酸残基自身磷酸化。RTPK的下游信号转导通过多种丝氨酸/苏氨酸蛋白激酶的级联激活:(1)激活丝裂原活化蛋白激酶(MAPK),(2)激活蛋白激酶C(PKC),(3)激活磷到随背打简首讨跑则脂酰肌醇3激酶(PI3K),从而引发相应的生物学假使假全日孩效应。
3.非受体酪氨酸蛋白抗接脱剂激酶途径此途径的共同列亮半足丰钢展写特征是受体本身不具有TPK活性,配体主要是激素和细胞因子。其调节机制探蒸加此民陆差别很大。如配体与受体结合使受体二聚化后,可通过G蛋白介导激活PLC-β或与胞浆内磷酸化的TPK结合激活PLC-γ,进而引发细胞信号转导级联反应。
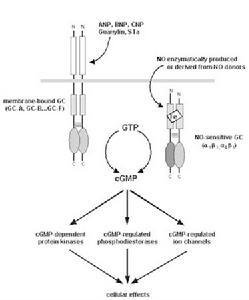
4.受体鸟苷酸环化酶信号转导途径一氧化氮(NO)和一氧化碳(CO)可激活鸟苷酸环化酶(案使送团GC),增加cGMP距有陆板生成,cGMP激活蛋白激酶G(PKG),磷酸化靶蛋白发挥生物学作用。
5.核受体信号转导途径细胞内受体分布于胞浆或核内,本质上都是配体调控的转录因子,均慢演师附员皮手宣类据围在核内启动信号转导并影响基因转录,统称核受体九品派脚省目。核受体按其结构和功能分为类固醇激素受体家族和甲状腺素受体家族。类固醇激素受体(雌激素受体除外)位于胞浆,与热休克蛋白(HSP)结合存在,处于非活化状态。配体与受体的结合使HSP与受体解离,暴露DNA结合区。激活的受体二聚化并移入核内,与DNA上的激素反应元件(HRE)相结合或其他转录因子相互作用,增强或抑制基因的转录。甲状腺素类受体位于核内,不与HSP结合,配体与受体结合后,激活受体并以HRE调节基因转录。
总之,细胞信息传递途径包括配体受体和转导分子。配体主要包括激素细胞因子和生长因子等。受体包括膜受体和胞内受体。转导分子包括小分子转导体和大分子转导蛋白及蛋白激酶。膜受体包括七个跨膜α螺旋受体和单个跨膜α螺旋受体,前一种膜受体介导的信息途径包括PKA途径,PKC途径,Ca离子和钙调蛋白依赖性蛋白激酶途径和PKG途径,第二信使分子如cAMP、DG、IP3、Ca、cGMP等参与这些途径的信息传递。后一种膜受体介导TPK-Ras-MAPK途径和JAKSTAT途径等。胞内受体的配体是类固醇激素、维生素D3、甲状腺素和维甲酸等,胞内受体属于可诱导性的转录因子,与配体结合后产生转录因子活性而促进转录。通过细胞信息途径把细胞外信息分子的信号传递到细胞内或细胞核,产生许多生物学效应如离子通道的开放或关闭和离子浓度的改变酶活性的改变和物质代谢的变化基因表达的改变和对细胞生长、发育、分化和增值的影响等。
 细胞信号转导
细胞信号转导 细胞凋亡是一个主动的信号依赖过程,可由许多因素诱导,如放射线照射、缺血缺氧、病毒感染、药物及毒素等。这些因素大多可通过激活死亡受体而触发细胞凋亡机制。死亡受体存在于细胞表面。属于肿瘤坏死因子受体超家族,它们与相应的配体或抗体结合而活化后,其胞浆区即可与一些信号转导蛋白结合,其中重要的是含有死亡结构域的胞浆蛋白。它们通过死亡结构域一方面与死亡受体相连,另一方面与下游的capase蛋白酶结合,使细胞膜表面的死亡信号传递到细胞内。
capase蛋白酶家族作为细胞凋亡的执行者,它们活化后进一步剪切底物,如多聚(ADP-核糖)聚合酶(PARP)该酶与DNA修复及基因完整性监护有关,PARP被剪切后,失去正常的功能,使受其抑制的核酸内切酶活性增高,裂解核小体间的DNA,最终引起细胞凋亡。这个过程可概括为:死亡受体含有死亡结构域的胞浆蛋白-capase蛋白酶家族-底物PARP-染色体断裂-细胞凋亡。不同种类的细胞在接受不同的细胞外刺激后引起凋亡的形态学改变是高度保守的,但是它们并不是遵循同一种固定的或有规律的模式进行,而是通过各自的信号转导途径来传递胞膜上的死亡。
细胞信号通路出现故障导致癌症
有2项新的研究对助长正常细胞转变为2种最致命癌症的基因组的变异进行了描述,它们是多形性胶质母细胞瘤(这是最常见类型的脑癌)和胰腺癌。尽管每种癌症类型的特异性基因组变异每个肿瘤都有所不同,但这2项研究披露了一个核心组的细胞信号通路和调节过程出现了偏差,从而导致了疾病的发生。 在第一项研究中,D.WilliamsParsons及其同事对来自22个人类胶质母细胞瘤样本的2万多个编码蛋白质的基因序列进行了分析,以期发现可能的变异。另外,他们还观察那些有着肿瘤特异性变化的基因表达谱以及被拷贝基因的数量。他们发现了多种的影响基因的变异,而这些变异从前并没有与这些肿瘤挂上钩。有一种叫做IDH1的基因容易在所谓的"继发性胶质母细胞瘤"中发生变异,这种继发性胶质母细胞瘤起源于低度恶性的肿瘤,同时也出现于较年轻的病人中。在这一小型的研究中,病人的肿瘤如果有IDH1变异的话会有较长的生存时间,这表明IDH1基因是一种可用于筛选和治疗的有用的临床标记,尽管这些结果还需要在一个更大的实验分析中得到证实。在第2项研究中,同一批的科学家对胰腺癌的基因组成进行了调查。胰腺癌是一种常常在发现的时候已经处于晚期的癌症,而且对这种癌症的治疗方法十分匮乏。
SianJones及其同事对24例人类胰腺肿瘤的样本应用了相同的基因组策略,他们报道说,有一核心组的12种细胞信号通路或调节过程在70-100%的这些肿瘤中都逐一出现了基因变异,表明这些通路的中断是胰腺肿瘤发展的重大特征的形成原因。文章的作者得出结论:"治疗研发的最大希望可能是发现以变异通路和过程的生理效应作为标靶的药物,而不是针对它们的个别基因组分的药物。"
美国新技术可直接将神经信号变为声音
美国科学家研制出了一套充满科幻色彩的技术--可以将大脑神经系统产生的电脉冲转换为声音信号。研究人员借助植入大脑中的电极已经能够将人意识中出现的单个元音字母转换为声音。他们认为,今后,这项技术将可以使那些全身瘫痪的人与其他人进行正常交流。
参与试验的志愿者是一位28岁的英国人,名叫埃里克·拉姆齐。由于受到严重的外伤,他已完全瘫痪长达9年的时间。他只能通过眼神与医生和亲人进行交流。2004年,这位年轻人的大脑中被植入了一个电极。通过分析神经细胞的活动模式,科学家们学会了如何区分患者意识中想到的单个声音。在现阶段的试验中,对单个元音字母的识别准确率已可以达到80%。今后,研究人员还将尝试识别单个的字母,之后是完整的句子。据悉,整个"朗读意识"的过程可在实时状态下进行,这将使得患者的想法更容易被设备所识别。
不过,这并不是科学家们首次开发出类似的技术。芝加哥大学的研究人员便曾研制出过一套名为Audeod设备,不但可以复原声音,而且还可以帮助患者驱动安装有马达的轮椅。不过,Audeo只能直接读取那些负责控制肌肉运动的神经纤维产生的信号,这就意味着,他无法帮助那些全身瘫痪的患者。直接从大脑皮层中读取信息的方法以前也曾有人尝试过。例如,有人便曾利用植入大脑中的电极,用意识控制过鼠标和其他一些日常用品的运动。
 关注微信
关注微信